Introduction
Humans are exposed to arsenic through food, drinking water, and soil (
Karagas et al. 1998). Arsenic poisoning is known to cause damage to several organs and tissues (
Kurttio et al. 1999). Chronic arsenic exposure has also been linked to a significantly higher risk of cancer of the lung, liver, bladder, kidney, and colon (
Smith et al. 2009). Many studies have shown that arsenic ingestion for a long period results in cutaneous lesions, hyperkeratosis, necrotic pigmentation, and melanosis (
Fatmi et al. 2009). Arsenic causes chromosomal aberrations, sister chromatid exchange, and the alteration of DNA-methylation (
Zhao et al. 1997). Long term exposure of arsenic in animal models has established male and female reproductive hazards: the alteration of ovarian and testicular steroidogenesis, the disruption of ovarian and testicular tissue, and the arrestation of spermatogenesis and folliculogenesis (
Shukla and Pandey 1984;
Chattopadhyay and Ghosh 2010). Arsenic-contaminated drinking water has xenoestrogenic effects on the anterior pituitary gland, and anterior pituitary hormone secretion is affected by arsenic (
Ronchetti et al. 2016). Chronic environmental arsenic exposure in humans has been reported to cause reproductive disturbances in women (
Zadorozhnaja et al. 2000) including critical pregnancy, low birth weight, reduced lactation, and impetuous abortion (
Ahmad et al. 2001). Sodium arsenite causes slow activity of the hypothalamic-pituitary-gonadal axis and suppresses natural folliculogenesis by downregulating the plasma follicle stimulating hormone (FSH) (
Chattopadhyay and Ghosh 2010). Arsenic ingestion for a long duration in humans elevates necrotic toxicity markers like lactate dehydrogenase (LDH) (
Karim et al. 2010).
The invasive arsenic chelators British anti-Lewisite (BAL) and dimercaptosulfonic acid (DMSA) are frequently used for the removal of arsenic from the body. However, long term use of this treatment strategy is problematic because of the complexity of the treatment procedure as well as the moderate to severe side effects of these chelators (
Inns et al. 1990). The detoxification of arsenic is a challenging issue, and investigators are now focusing on the use of different antioxidants, natural products, and plant components. All-trans retinoic acid (ATRA) relieved sodium-arsenite-induced reactive oxygen species (ROS) generation, restored redox balance, and prevented apoptosis (
Chatterjee and Chatterji 2017). Vitamin E and selenium conjointly or alone ameliorated arsenic-induced cardiotoxicity (
Bhattacharjee and Pal 2014) and female reprotoxicity (
Chattopadhyay et al. 2000).
Recently, a microscopic blue-green alga (cyanobacterium), spirulina (
Spirulina platensis (
Geitler 1925)), is showing evidence of therapeutic importance, as it appears to reduce nephrotoxic substances in the body (
Khan et al. 2006). Phycocyanin and the natural carotene in spirulina could potentially inhibit cancer cell development and growth (
Peto et al. 1981;
Shekelle et al. 1981). C-phycocyanin was reported as a potentially allergenic protein (
Le et al. 2014) in spirulina-mediated anaphylaxis; however, preclinical studies related to acute and chronic toxicity, teratogenicity, fertility, and reproductive function assessments have demonstrated that spirulina itself is not toxic (
Chamorro et al. 2002). Spirulina has also been listed as “generally recognized as safe” by the US Food and Drug Administration (
Belay 2008).
Studies have shown that the antioxidant and anti-inflammatory effects of spirulina assist in the treatment of diabetes, hypertension, and hyperlipidaemia (
Lee et al. 2008). Spirulina effectively promotes cell rejuvenation and, thereby, repairs liver and kidney tissue damage (
Khan et al. 2001). In addition, spirulina has a protective effect against arsenic toxicity in humans (
Misbahuddin et al. 2006;
Rahman et al. 2008). Therefore, the aim of our investigation was to elucidate the role of the non-invasive oral application of spirulina in the amelioration of high dose arsenic-mediated uterine degeneration.
Materials and methods
Animal selection and care
In this experimental study, adult female albino rats (80–100 g body mass) were purchased from the approved animal provider Saha Enterprise, Kolkata, West Bengal. We conducted this study on Wistar strain adult female albino rats. The study was conducted with the approval of the Vidyasagar University’s ethical committee (ethical clearance No. IEC/7-2/C-2/16, dated 26 August 2016) and all experimental procedures were done according to the standards of The Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), India guidelines. The rats were reared in polycarbonate cages in the institutional animal house. They were adapted for 10 d in a room with a 12 h light:dark cycle, with 32 ± 2 °C temperatures and 50%–70% humidity. A standard pellet diet (Hindustan Lever Ltd., Mumbai, India) and water ad libitum were provided to the rats.
Treatment, study of estrous cycle, and sample collection
Twenty-four rats were equally and randomly separated into four groups of six. The vehicle-treated control group received only distilled water, the arsenic-treated group received 1.0 mg of sodium arsenite/100 g body mass, the spirulina placebo group received 20 mg of spirulina/100 g body mass, and the arsenic + spirulina treatment group received both 1.0 mg of sodium arsenite/100 g body mass and 20 mg spirulina/100 g body mass. In this experiment, 1.0 mg of sodium arsenite was dissolved in 0.1 mL of distilled water and applied per 100 g body mass of the rats. Spirulina tablets (500 mg, West-Coast Pharmaceutical, Ahmedabad, Gujarat) were crushed to produce spirulina dust, and 20 mg of spirulina were dissolved in 0.15 mL of sterile distilled water per 100 g body mass of the rats fresh daily before each use. Rats were treated via oral gavage for 8 d. The body masses of all rats were recorded throughout the treatment. The pattern of the estrous cycle was observed. Vaginal smear was collected daily from all animals. The feeding habits of all rats were observed throughout the treatment. On day nine the final body masses were recorded. Wistar rats were anesthetized with ketamine, and organs were collected from the anesthetized Wistar female rats following the standard protocol of Vidyasagar University’s ethical guidelines (ethical clearance No. IEC/7-2/C-2/16, dated 26 August 2016). Blood was collected for the separation of serum and then the animals were euthanized with CO2 as per the CPSCEA guideline. Dissected organs were preserved at − 20 °C until use.
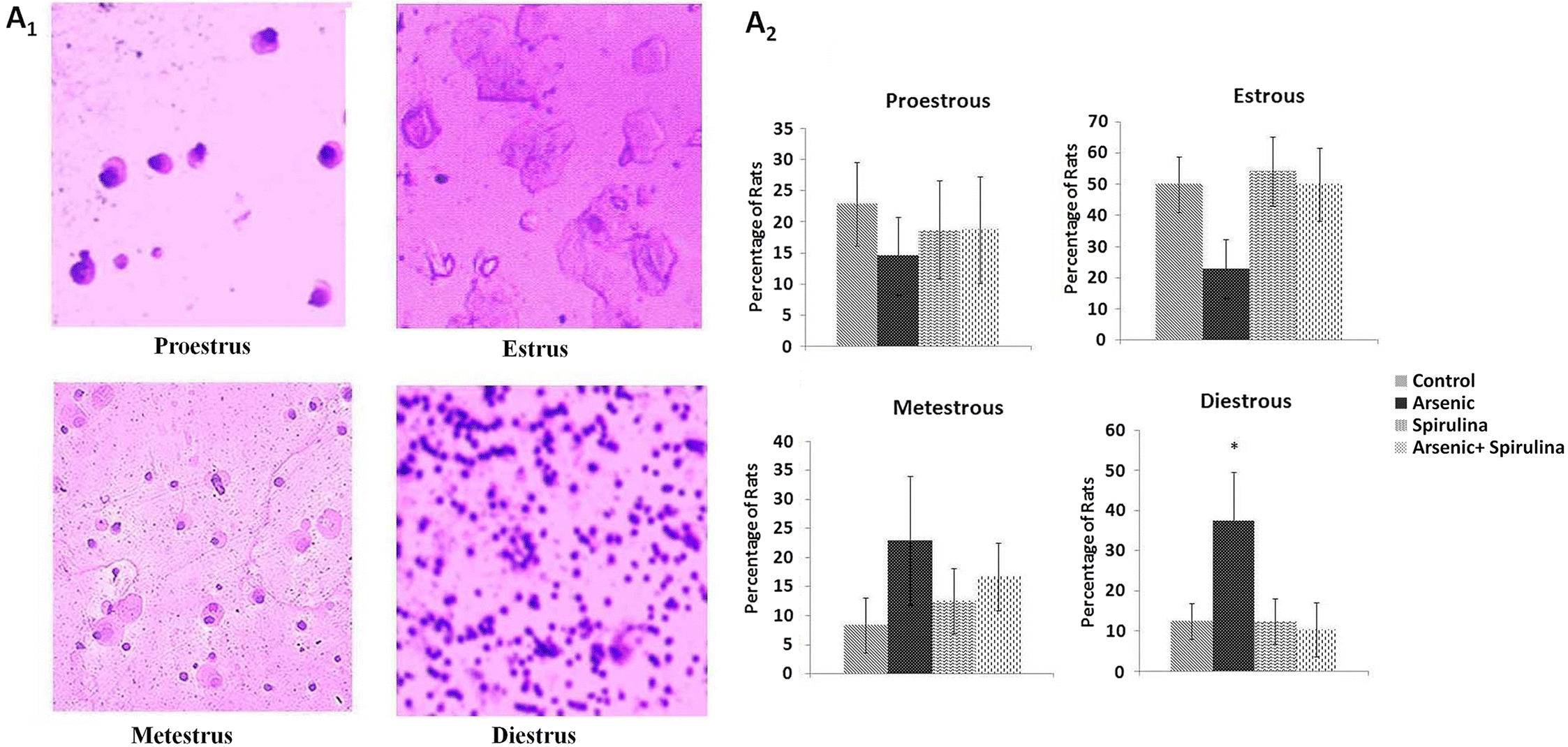
Study of the estrous cycle
The different stages of the estrous cycle were studied in all rats following the collection of vaginal smears. The collected vaginal fluid was uniformly spread on a glass slide. The glass slides were allowed to dry for a few minutes and then were stained with Leishman stain and the phase of the estrous cycle was identified using a microscope (100× magnification).
Estimation of malondialdehyde (MDA) and conjugated dienes (CD) levels
Uterine tissues were homogenized (20% w/v) in ice-cold phosphate buffer (0.1 mol/L, pH 7.4) and centrifuged at 15 000g at 4 °C for 3 min. Supernatant was collected for the estimation of MDA and CD.
MDA levels were determined from the reaction of thiobarbituric acid with MDA. The amount of MDA formed was measured (
Devasagayam et al. 2003) by taking the absorbance at 530 nm (
ε = 1.56 × 10
5 L/mol·cm).
Conjugated dienes levels were determined using a standard method. The lipids were extracted with chloroform–methanol (2:1) and centrifuged at 1000
g for 5 min. The residue of the lipid was then dissolved in 1.5 mL of cyclohexane. The hydroperoxide formed was measured at 233 nm (
Kumar 2012).
Assay of super oxide dismutase (SOD), catalase, peroxidase, and lactate dehydrogenase (LDH) activities
Uterine tissues were homogenized in 100 mmol/L Tris–HCl (pH 7.4) to make a tissue concentration of 10%
w/
v and centrifuged at 10 000
g for 20 min at 4 °C. A 100 μL portion of the tissue supernatant was mixed with 800 μL of triethanolamine-diethanolamine–HCl buffer (TDB) (Merck & Company, Inc., Kenilworth, New Jersey, USA), 40 μL of 7.5 mmol/L nicotinamide adenine dinucleotide phosphate (NADPH) (Sigma-Aldrich Corporation, St. Louis, Missouri, USA), and 25 μL of ethylenediaminetetraacetic acid (EDTA)-MnCl
2 (Himedia, Mumbai, India). The activity of SOD was monitored at 340 nm from the rate of oxidation of the NADPH (
Pattichis et al. 1994).
The activity of the catalase was measured colorimetrically. Uterine tissue was homogenized with ice-cold phosphate-buffered saline (PBS) (0.1 mol/L, pH 7.4) and centrifuged at 10 000
g for 20 min at 4 °C. Here, dichromate in acetic acid was transformed first into perchromic acid and then into chromic acetate when heated in the presence of H
2O
2. The resulting chromic acetate was measured at 570 nm (
Hadwan 2016). The catalase preparation permitted the splitting of H
2O
2 for different durations. The reaction was terminated at different time intervals by the addition of a dichromate–acetic acid mixture. One unit of activity was identified as one mole of H
2O
2 consumed per minute per milligram of protein.
For the peroxidase assay, uterine tissue was homogenized using PBS (0.1 mol/L, pH 7.0). A cocktail of 20 mmol/L of guiacol and 0.1 mL of supernatant was used in the presence of H
2O
2 (12.3 mmol/L) as a substrate. The time when the absorbance (436 nm) increased by 0.1 unit (
Sadasivan and Manickam 1966) was noted.
The LDH serum levels were measured using LDH (P-L) kits, following the manufacturer’s (Tulip Group, Goa, India) instructions. LDH reduces pyruvate in the presence of NADH and thereby forms NAD. The rate of oxidation of NADH to NAD was determined by a decrease in the absorbance, which was proportional to the LDH activity.
Assessment of SOD, catalase, peroxidase, and LDH in native gel
Uterine tissues were homogenized with ice-cold PBS (1.0 mol/L, pH 7.4) and centrifuged at 10 000
g for 20 min at 4 °C. The 40 μg of proteins from the tissue extract were applied to 12% polyacrylamide gel electrophoresis (PAGE). The gel was incubated with 2.3 mmol/L nitroblue tetrazolium (NBT), 28 μmol/L riboflavin, and 28 mmol/L tetramethylethylenediamine (TEMED) for 20 min in dark conditions. Upon light exposure the achromatic bands of SOD were visualized against a dark blue background (
Weydert and Cullen 2010).
Uterine tissue extract containing 50 μg of proteins was electrophoresed on 8% PAGE for catalase detection. Gels were kept in 0.003% H
2O
2 solution for 10 min and stained with 2% potassium ferricyanide and 2% ferric chloride. Bluish-yellow bands appeared against a blue and green background (
Lewis et al. 2005).
The 50 μg of proteins present in the tissue extract were loaded in 8% native gel and the gel was run with a power supply of 40 mA. The gel was then stained with 100 mg of benzidine powder in the presence of 4.5 mL glacial acetic acid and 30% H
2O
2 to observe the peroxidase band (
Hasan and Aburahma 2014).
Electrozymography of LDH was performed in 8% agarose gel in 50 mmol/L Tris–HCl (pH 8.2) and 20 μL of serum were loaded into the different gel slots. The gel was electrophoresed at 170 V. The gel was incubated at 37 °C in the presence of Na-lactate, NAD and 1.0 mol/L Tris to develop the colour reaction in association with NBT and phenazine methosulphate. Finally, the gel was rinsed with water under light (
Brandt et al. 1987).
DNA fragmentation analysis
Uterine tissues were used for DNA preparation and treated with 500 μL lysis buffer (50 mmol/L Tris-pH 8.0, 20 mmol/L EDTA, 10 mmol/L NaCl, 1% SDS, 0.5 mg/mL proteinase K) for 15 min at 4 °C. Centrifugation was performed at 12 000 rpm for 20 min. The collected supernatant was added to a 1:1 mixture of phenol:chloroform, gently agitated for 5 min, then precipitated into two parts of cold ethanol and one tenth part of sodium acetate (
Paoletti et al. 1990). After spinning down and decantation, the precipitate was resuspended in 30 μL of deionized water–RNase solution (0.4 mL water + 5 μL of RNase) and 5 μL of loading buffer for 30 min at 37 °C. The 8.0% agarose gel with ethidium bromide was electrophoresed at 65 V and documented in a gel documentation system (
Garcia-Martinez et al. 1993).
Comet assay
Slides were dipped into a 1% hot agarose solution. The uterine cell suspension was exposed to low melting point 0.6% agarose in PBS at 37 °C, fixed onto a glass slide precoated with 1% agarose, and a cover-slip was placed on the slide. A slight modification of the method described by
Singh et al. (1988) was followed for the comet assay. Electrophoresis was performed for 30 min at 25 V and the current was adjusted to 300 mA by raising the buffer level. Slides were then neutralized with PBS and stained with a solution of 10 mg/mL ethidium bromide for 5 min. Excess stain was removed by washing with water. The slides were observed under a fluorescence microscope (Eclipse LV100 POL, Nikon, Tokyo, Japan) using Vis Comet software (Impuls Bildanalyse, Amsterdam, The Netherlands).
Serum hormone analysis
Serum levels of leutinizing hormone (LH) (Cat. No. ER1123), follicle stimulating hormone (FSH) (Cat. No. ER0960), and estradiol (Cat. No. ER1507) were measured using enzyme-linked immunosorbent assay (ELISA) kits following the procedures recommended by the manufacturer (Wuhan Fine Test, China).
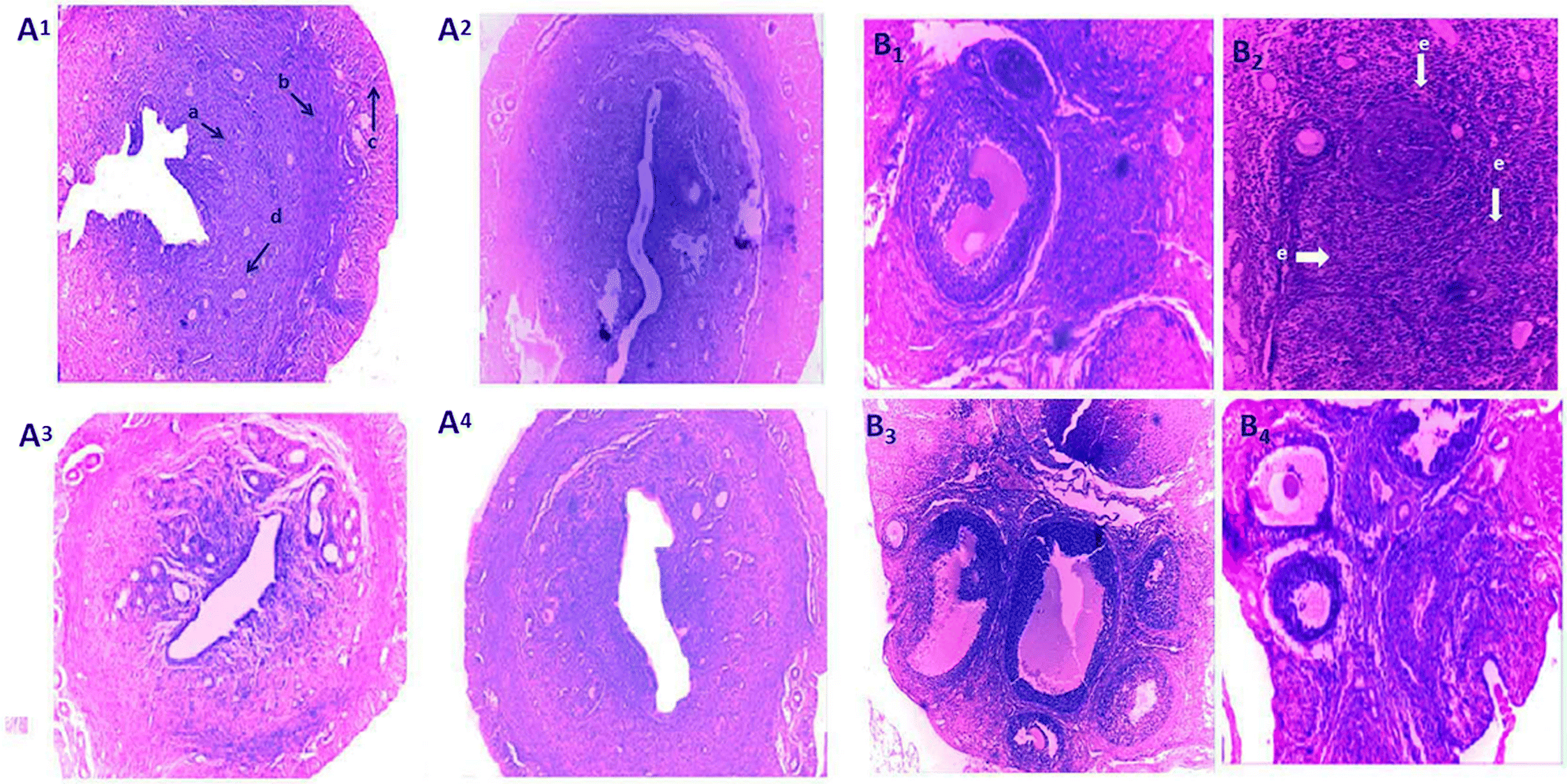
Histo-architecture of ovary and uterus
Ovarian and uterine tissues were embedded in paraffin and sectioned into 5 μm slices, and stained with hematoxylin (Harris) and eosin (HE). Stained samples were observed under a microscope (CX21i, magnification 100× , Olympus, Tokyo, Japan).
Statistical analysis
In this experiment, data were expressed as mean ± SE, with n = 6 for each of the four experimental treatment groups (N = 24). Data from the supplemented group were compared with the control by analysis of variance (ANOVA) followed by post-hoc Dunnett’s t test and post-hoc least significant difference (LSD) test. p < 0.05 was considered a minimum level of significance.
Discussion
A trivalent form of arsenic (As
III) is recognized for its high degree of toxicity. Previously, a comparatively low dose of arsenic for 28 d (
Chattopadhyay et al. 2012;
Acharyya et al. 2015) was considered to demonstrate the toxic effect of arsenic on different body parts. Our study used a comparatively higher dose of arsenic (10 mg/kg body mass) with 8 d of treatment to evaluate the effects of arsenic toxicity in female reproductive organs. Our study also focused on whether spirulina is effective against such a high dose of arsenic. However, arsenic at a dose of 10 mg/kg body mass for a short duration exhibited a significant level of free radical generation in the organs (
Manna et al. 2007;
Sinha et al. 2008;
Bhattacharya and Haldar 2013). Arsenic treatment at a dose of 10 mg/kg body mass for a short duration like 8 d was previously used by our group (
Maity et al. 2018). In this experiment, arsenic (As
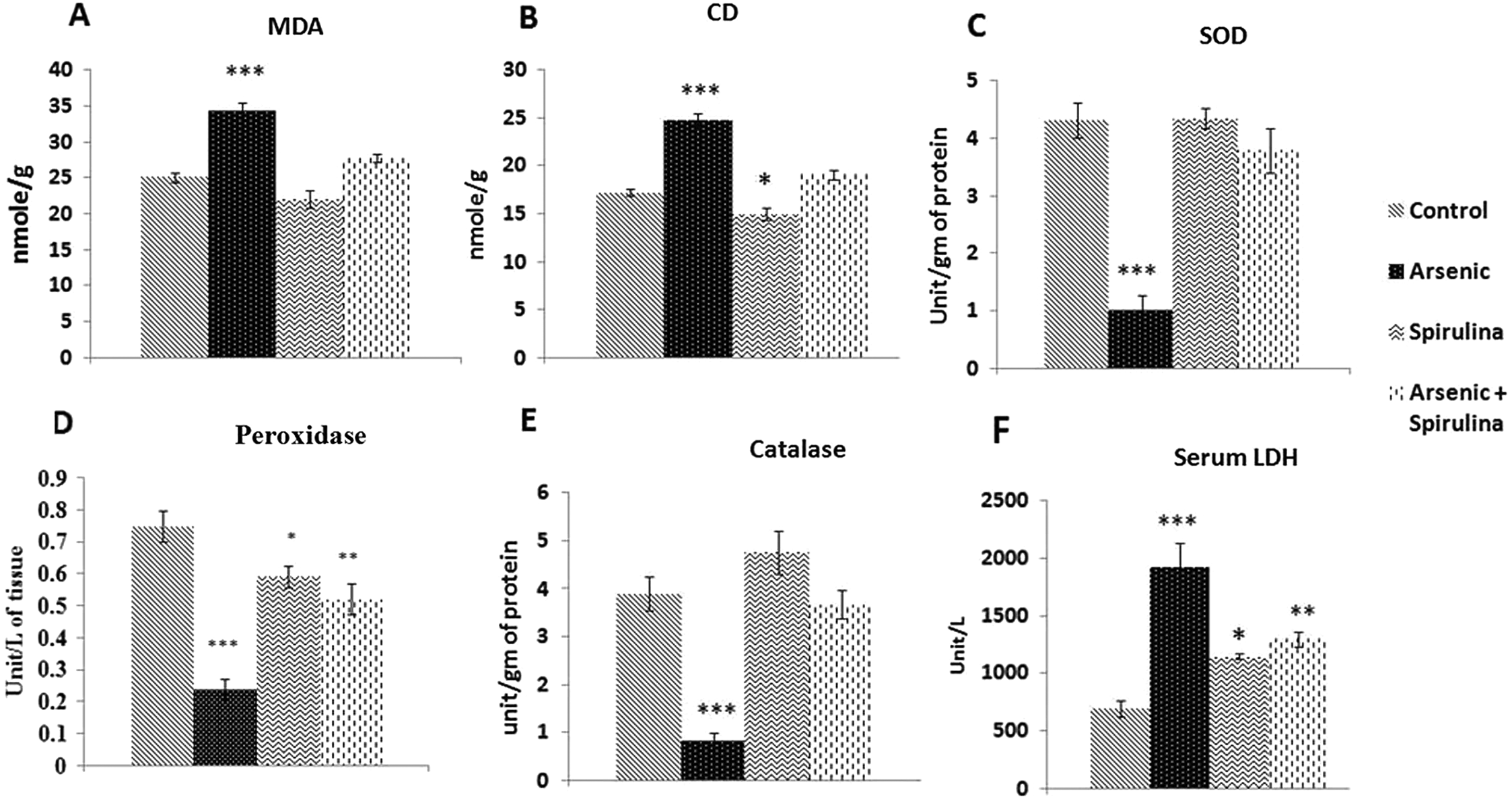
III) was involved in the generation of oxidative stress and manifested in elevated uterine MDA and CD levels (
Fig. 2). This indicates that the trivalent form of arsenic yields reactive oxygen species in association with H
2O
2 and, therefore, produces end products in the form of lipid peroxides and conjugated dienes (
Wang and Xu 2006). Electrozymograms of SOD in our investigation revealed a decreased intensity of SOD bands in the arsenic-treated group (
Fig. 3A). The activity of SOD in the gel was shown to be suppressed by arsenic after staining with NBT.
Rana et al. (2012) also documented that arsenic reduced mRNA expression of the SOD gene. Furthermore, in the current study we electrozymographically documented decreased catalase and peroxidase activity indicated by the existence of weak bands following arsenic ingestion (
Figs. 3B and
3C). These results indicate the possible impairment of H
2O
2 detoxification in the uterine tissue due to the diminished activity of catalase. Previous studies have shown that the inhibition of catalase by arsenic trioxide resulted in intracellular ROS accumulation (
Wang et al. 2012). This impairment in catalase activity by arsenic may be mediated via the modulation of its expression at the level of mRNA transcription (
Wang et al. 2012). Likewise, our result revealed that the expression of peroxidase was also reduced in response to arsenic. This finding strongly suggests that H
2O
2 accumulates during programmed cell death (
Weydert and Cullen 2010).
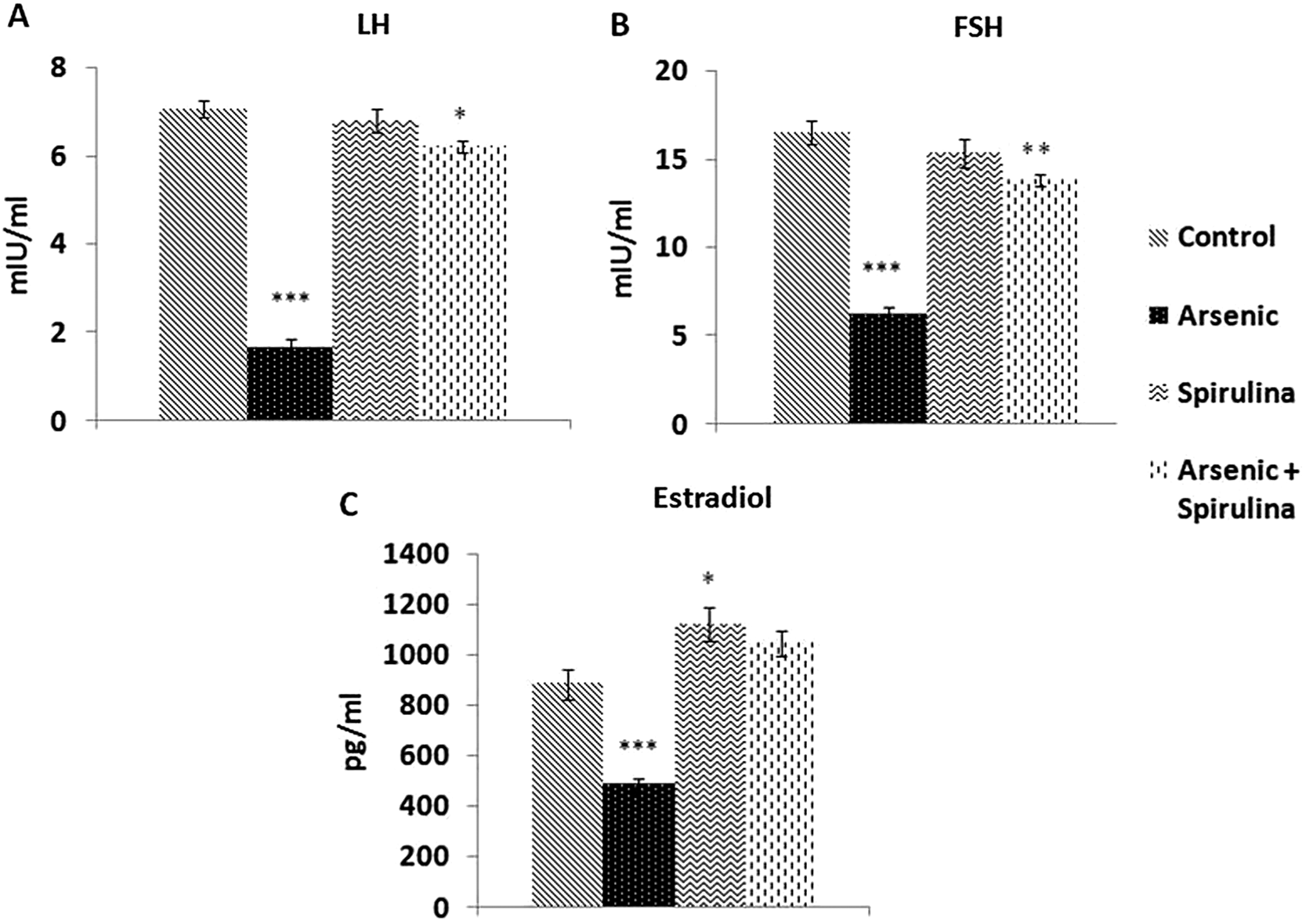
Arsenic exposure had detrimental effects on plasma levels of LH, FSH, and estradiol (
Fig. 4) along with low ovarian and uterine mass (
Fig. 4). Rats remained in consistent diestrus or metestrus after 4 d of treatment compared with the control rats, whereas synchronization of the estrous cycle was achieved following the treatment with spirulina. Consistent diestrus in arsenic-treated rats after 5 d may be due to low estrogen plasma levels (
Parshad et al. 1989). This indicates that arsenic indirectly imposes its effect on ovaries by altering the regulation of the pituitary–ovarian axis. It is possible that the deregulation of the gonadotrophins from arsenic may result in low ovarian mass and follicular regression (
Tagatz et al. 1970;
Kulin and Reiter 1973), whereas inhibition of the estradiol signalling in these rats may be the limiting factor causing decreased uterine mass and the degradation of the normal uterine histo-architecture (
Edman 1983).
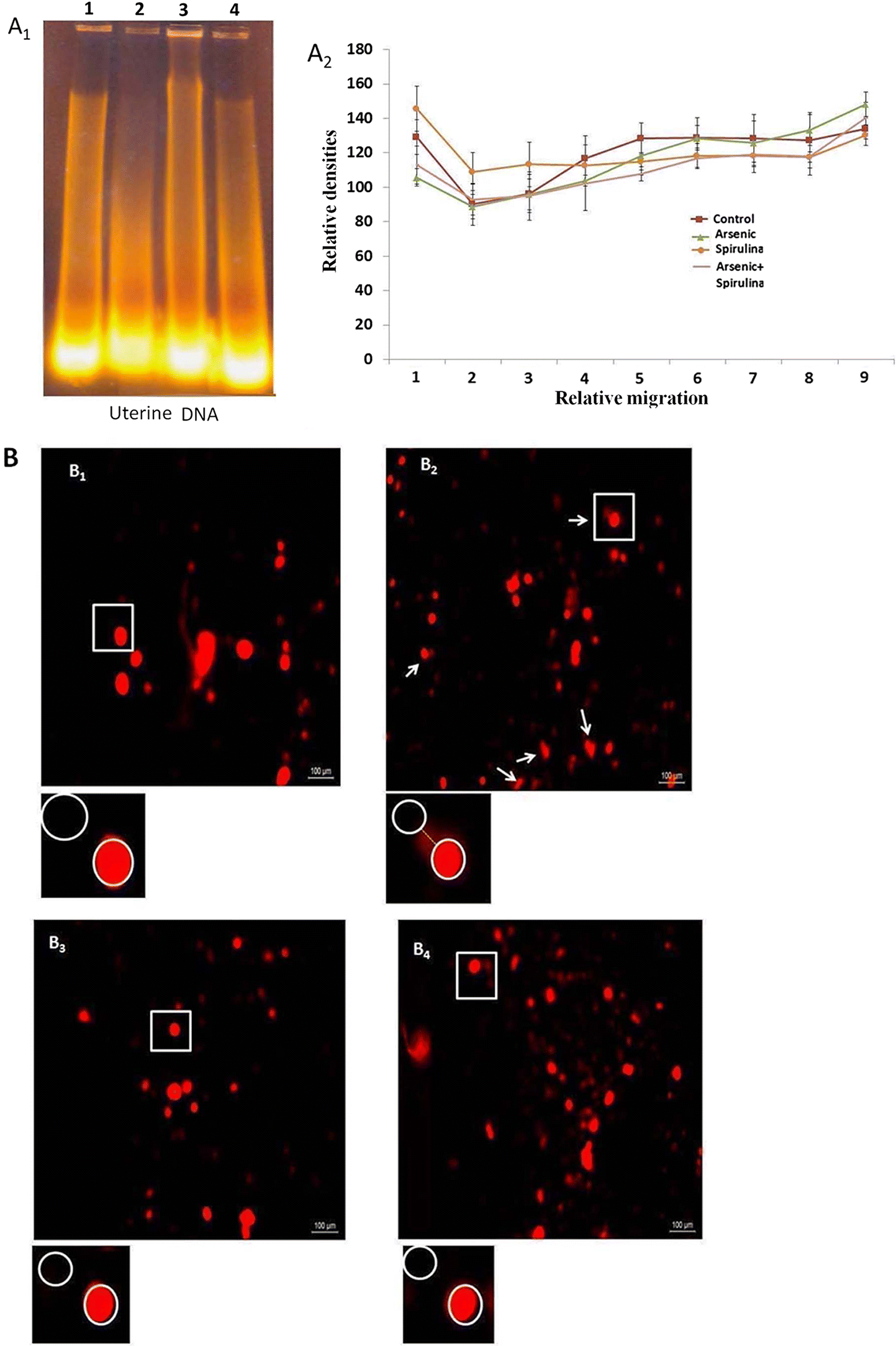
Spirulina co-treatment significantly mitigated arsenic-induced uterine oxidative stress and ovarian disorders. Spirulina is a non-enzymatic antioxidant and may, thereby, improve the enzymatic antioxidant status. The cellular degeneration from arsenic exposure was likely necrotic in nature, which can be demonstrated by uterine DNA fragmentation (
Fig. 5A1). In this study, arsenic showed conspicuous DNA fragmentation (
Fig. 5A1 Lane 2) in the uterine tissues compared with the control (
Fig. 5A1 Lane 1). The formation of ROS by trivalent arsenicals may ultimately lead to DNA damage, thereby initiating a suppression of DNA repair systems and the repair of oxidative DNA damage (
Kligerman et al. 2003). In this experiment, the apoptotic features were evident from the comet assay and DNA fragmentation image (
Figs. 5B and
5A1). This study demonstrated that co-treatment with arsenic + spirulina (
Fig. 5A1 Lane 3, 4) partially, but significantly, reduced the degradation of DNA caused by the arsenic. The protection of DNA by spirulina against arsenite-induced toxicity was also demonstrated in single cell DNA damage (
Fig. 5B;
Table 2). A low serum LDH and the protection of DNA damage following arsenic + spirulina co-administration in rats may act as the barrier to necrotic progression (
Ding et al. 2005). This overall defensive mechanism of spirulina might have terminated arsenic-primed inducement of apoptosis and protected uterine and ovarian tissues from necrotic damage, as there was an improvement in the ovarian–uterine histo-architecture (
Figs. 6A and
6B). This may be connected with the upregulation of gonadotrophin release and ovarian steroidogenesis (
Tagatz et al. 1970;
Kulin and Reiter 1973).
Other investigators have explored the therapeutic effects of spirulina on arsenic-induced male reproductive toxicity (
Bashandy et al. 2015). Spirulina is a unique alga because it is enriched in essential amino acids such as sulfolipids, linolenic acid, protein, essential fatty acids, vitamins, flavonoids, and minerals (
Mendes et al. 2003;
Dartsch 2008). In addition, spirulina contains phycocyanin and phycocyanobilin (
McCarty 2013), which have been claimed to inhibit peroxyl-radical-induced lipid peroxidation and NADPH-dependent superoxide production (
Bhat and Madyastha 2001;
Zheng et al. 2013). Because it has the capacity to bind with heavy metals, spirulina may demonstrate a chelating property to trap arsenic (
Plazinski 2013). Methylation of arsenic in association with S-adenosylmethionine involves methyl cobalamin (CH
3B
12) and reduced glutathione for arsenic clearance and detoxification with the involvement of enzymes (
Nakamura 2011). Arsenite S-adenosyl methionine methyltransferase was isolated and characterized from spirulina (
Guo et al. 2016). An in vitro assay system demonstrated the arsenic methylation activity of spirulina (
Guo et al. 2016). This methylation process is the essential prerequisite for the removal of arsenic. Therefore, spirulina may eventually promote arsenic methylation and detoxification.