Ankylosaurus magniventris is an iconic dinosaur species often depicted in popular media. It is known from relatively fragmentary remains compared with its earlier and smaller relatives such as Euoplocephalus and Anodontosaurus. Nevertheless, the known fossils of Ankylosaurus indicate that it had diverged significantly in cranial and postcranial anatomy compared with other Laramidian ankylosaurines. In particular, the dentition, narial region, tail club, and overall body size differ substantially from other Campanian–Maastrichtian ankylosaurines. We review the anatomy of this unusual ankylosaur using data from historic and newly identified material and discuss its palaeoecological implications.
Introduction
The iconic Maastrichtian dinosaur Ankylosaurus magniventris was the last and largest of the ankylosaurid dinosaurs. A member of the tail-clubbed clade of armoured dinosaurs, the ankylosaurines, Ankylosaurus was part of a lineage that dispersed into North America from Asia during the Late Cretaceous (Arbour and Currie 2016). Like other ankylosaurines, Ankylosaurus was a bulky, broad quadruped studded with osteoderms of various shapes and sizes, and had a stiff distal tail with enlarged osteoderms enveloping the tail tip to form a formidable tail club. It was part of the charismatic dinosaur megafauna of the latest Cretaceous of western North America, living alongside Tyrannosaurus, Triceratops, and Edmontosaurus (Horner et al. 2011).
Despite its household name status, Ankylosaurus is known from far fewer remains than its Campanian–Maastrichtian relatives Euoplocephalus and Anodontosaurus, with only a handful of specimens from the Western Interior currently referable to this genus (Fig. 1). Nevertheless, these remains indicate that Ankylosaurus had diverged from the Laramidian ankylosaurine bauplan to a surprising degree, particularly with regards to its narial anatomy and body size. Carpenter (2004) was the last to review Ankylosaurus anatomy in detail, but a bevy of recent ankylosaurid finds (e.g., Loewen et al. 2013; Arbour et al. 2014a; Arbour and Evans 2017), new insights into ankylosaur anatomy and relationships (Witmer and Ridgely 2008; Thompson et al. 2012; Hill et al. 2015; Leahey et al. 2015; Arbour and Currie 2016), and novel information gleaned from historical specimens warrant a re-evaluation of the genus that we present here.
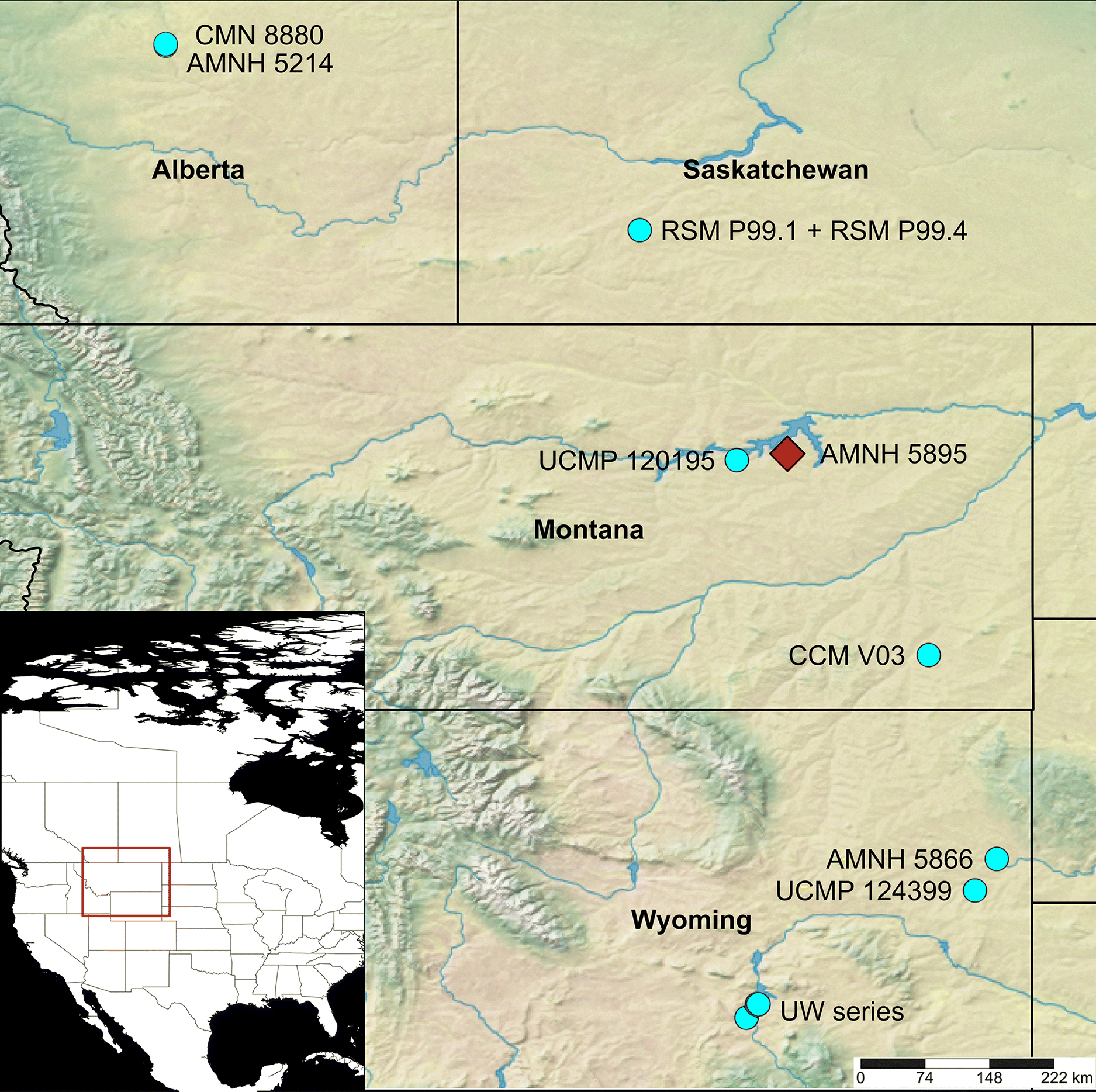
Fig. 1.
Fig. 1. Distribution map of known Ankylosaurus magniventris fossils. The red diamond marks the location of AMNH 5895, the holotype of the genus. Map generated using SimpleMappr (simplemappr.net).
We offer new observations concerning the cranial and postcranial anatomy of Ankylosaurus not already presented by Carpenter (2004). We describe previously unidentified and undescribed elements of the holotype, AMNH 5895, and of the largest specimen, CMN 8880. We also reinterpret previously established aspects of the anatomy of Ankylosaurus, including a new restoration of the arrangement of osteoderms. We conclude by providing some updated body size estimates for Ankylosaurus and tease out some of the implications regarding the palaeoecology of the species.
Materials and methods
Ankylosaurus is known from only a handful of specimens: AMNH 5895, the holotype, including a partial skull and the most complete postcranium of any of the known specimens; AMNH 5214, the best preserved skull and only well-preserved tail club; CMN 8880, the largest skull; CCM V03, a fragmentary tail club handle; AMNH 5866, osteoderms associated with “Dynamosaurus” (=Tyrannosaurus; Osborn 1905); and several isolated osteoderms (RSM P99.1, RSM P99.4; Burns 2009). Isolated teeth and osteoderms have also been recovered from Lancian-aged formations (Carpenter 1982a; Lillegraven and Eberle 1999) and are provisionally referred to Ankylosaurus based on their stratigraphic provenance and general similarity to those of more complete Ankylosaurus specimens.
Several trays of bone fragments are available for the holotype of Ankylosaurus, AMNH 5895. A 2015 review of this material by the first author resulted in the identification of multiple fragmentary pieces of the skull, armour, and pelvis for this specimen, which have not previously been described in the literature. The best preserved skull of Ankylosaurus, AMNH 5214, is mounted behind a glass panel that cannot be removed. A high-quality cast of this skull, made available by Research Casting International, is accessioned at the University of Alberta Laboratory for Vertebrate Paleontology (UALVP 54722) and was used as an additional reference for this specimen.
CMN 8880 was briefly described and figured by Carpenter (2004), who suggested that the skull roof was poorly preserved. However, the skull was stored upside down at the time so that the dorsal surface may have been unavailable for examination. The skull was flipped in 2014 for conservation and study of the dorsal surface. The skull roof is, in fact, in a fine state of preservation. The right lateral surface of CMN 8880 is crushed medially and the anterior portions of the premaxillae are broken, but the skull is otherwise complete and uncrushed. The dorsal surface is described and figured here for the first time (Fig. 2, Supplementary Material 1).
Fig. 2.
Fig. 2. Skull of CMN 8880, Ankylosaurus magniventris, in (A) dorsal, (B) ventral, (C) left lateral, and (D) right lateral views. The skull is well preserved on the dorsal and left lateral surfaces. The right lateral surface has caved inwards slightly, and the premaxillary beak is missing. Left lower jaw in (E) lateral and (F) medial view. (G) Dentary tooth in labial view.
The lower jaw of CMN 8880 was digitized in three dimensions using a GO!Scan 20 portable 3D scanner and the cranium was digitized using a GO!SCAN 50 (Creaform Inc., Lévis, Québec, Canada). The initial scans were acquired and processed in VXelements 6.0 VXscan software (Creaform Inc., Lévis, Québec, Canada). Cranial data were collected in 14 passes, with the scanner set to the maximum resolution of 0.500 mm. The first set of seven passes consisted of scans of the dorsal surface. The second set of seven passes consisted of scans of the ventral surface. Data were collected for the lower jaw in six passes consisting of two scans of both the lateral and medial surfaces, with the scanner set to a resolution of 0.200 mm. The “Use Natural Features” function was selected under “Positioning Parameters” for target acquisition. All scans were aligned using manual alignment, and then merged into a single watertight mesh using the VXelements 6.0 VXmodel software. Both models were reduced to 5 million faces each using MeshLab (64 bit) v. 1.3.4 beta (Cignoni et al. 2008).
Measurements were taken directly from specimens or casts using tape measures and digital calipers. The tail club of AMNH 5214 is inaccessibly mounted at an angle behind glass, but some length measurements were acquired using a measuring tape laid flush with the glass and a laser pointer squared to the glass. Other measurements for this tail club were estimated using ImageJ (Rasband 2017) and figures in Carpenter (2004) cross-referenced with measurements of the mounted specimen.
We follow the terminology proposed by Arbour and Currie (2013a) for discussing cranial ornamentation in ankylosaurids. The tile-like ornamentations across the skull of ankylosaurids are referred to as caputegulae (Blows 2001) and include a location modifier; e.g., prefrontal caputegulae are located on the prefrontal, frontonasal caputegulae are located on the frontals and nasals (where the boundary between the frontal and nasal is unclear), and so on. Cranial scalation patterns are often used as diagnostic characteristics for extant squamates, and scale shapes and patterns can be highly conserved within species (e.g., Dixon 2000; Zug 2013). In ankylosaurs, individuals within a single species have relatively conserved caputegulum patterns (with the greatest variation typically found in the exact number and arrangement of the frontonasal caputegulae), and many caputegulae are present in similar locations across multiple clades, suggesting they are homologous and correspond to underlying cranial architecture (Arbour and Currie 2013a; Arbour et al. 2014a; Arbour et al. 2014b; Arbour and Currie 2016).
Holotype: AMNH 5895—partial skull, two teeth, five cervical vertebrae, eleven dorsal vertebrae, three caudal vertebrae, right scapulocoracoid, ribs, osteoderms (including portions of both cervical half rings); newly identified material includes an otic capsule, maxilla fragment, right jugal, left jugal and quadratojugal, two sacral centra, and additional fragments of the cervical half rings.
Holotype locality and age: Gilbert Creek, somewhere in Section 27 or 28, Township 22N, Range 40E, Garfield County, Montana, USA (Fig. 1); Hell Creek Formation, 61–67 m below Cretaceous–Palaeogene boundary, late Maastrichtian (Carpenter 2004).
Referred specimens: AMNH 5214—complete skull, both mandibles, six ribs, seven caudal vertebrae including tail club, both humeri, left ischium, left femur, right fibula, osteoderms (left bank of Red Deer River, centre of S26, T33, R22, Alberta, Canada; Scollard Formation, 45.4 m below K–Pg boundary, Maastrichtian; Sternberg 1951; Carpenter 2004). AMNH 5866—more than 70 osteoderms (Seven Mile Creek drainage, S14–16, T40N, R63W, Niobrara County, Wyoming, USA; Lance Formation, Maastrichtian, Carpenter 2004). CCM V03—partial tail club handle (east side along Powder River drainage, somewhere in R1S, T53E, about 24 km south of Powderville, Powder River County, Montana, USA; upper Hell Creek Formation; Maastrichtian; Carpenter 2004). CMN 8880—skull and left mandible (right bank of Red Deer River, SE ¼ S35, T33, R22, Alberta, Canada, 43.9 m below K–Pg boundary; Maastrichtian; Carpenter 2004). RSM P99.1 and RSM P99.4—osteoderms (coulee south of the village of Simmie, Saskatchewan; Frenchman Formation; Maastrichtian; Burns 2009). UCMP 120195—tooth (UCMP locality V-73076, Garfield County, Montana; Hell Creek Formation, Maastrichtian; Carpenter 1982a). UCMP 124399—tooth (UCMP locality V-5620, Niobrara County, Wyoming; Lance Formation; Maastrichtian; Carpenter 1982a). UW 26291 and UW 26293—teeth, and UW 26294 and UW 27418—osteoderms (UW localities V-93002, V-93016, V-92027, V-92041, and V-92012, Albany County, Wyoming; Ferris Formation, Maastrichtian; Lillegraven and Eberle 1999) (Fig. 1).
Revised Diagnosis: Ankylosaurine ankylosaur with a pattern of flat, hexagonal frontonasal caputegulae. Uniquely among ankylosaurids, nasal vestibule roofed entirely by loreal caputegulum and not supranarial caputegulum as in Anodontosaurus, Euoplocephalus, Scolosaurus, Ziapelta, and Zuul; the nasal vestibule is partially roofed by the loreal and supranarial caputegulae in Nodocephalosaurus. External nares open ventrolaterally; narial opening is not visible in anterior view. Loreal caputegulum is laterally expanded and bulbous. Keel of anterior and posterior supraorbital caputegulae is continuous with keel of squamosal horn. 34–36 proportionally small maxillary teeth (<2% basal skull length); greater number of maxillary teeth than in Anodontosaurus or Euoplocephalus. Neural spines of tail club handle vertebrae are U-shaped in dorsal view (not V-shaped as in other ankylosaurids). Compared with the handle vertebrae of Anodontosaurus and Euoplocephalus of the same length, the neural arch of the handle vertebrae in Ankylosaurus is at least twice as wide. Tail club knob approximately as transversely wide as anteroposteriorly long (also present in Euoplocephalus and Scolosaurus), not wider than long as in Anodontosaurus or longer than wide as in Dyoplosaurus.
New anatomical observations
Cranial anatomy
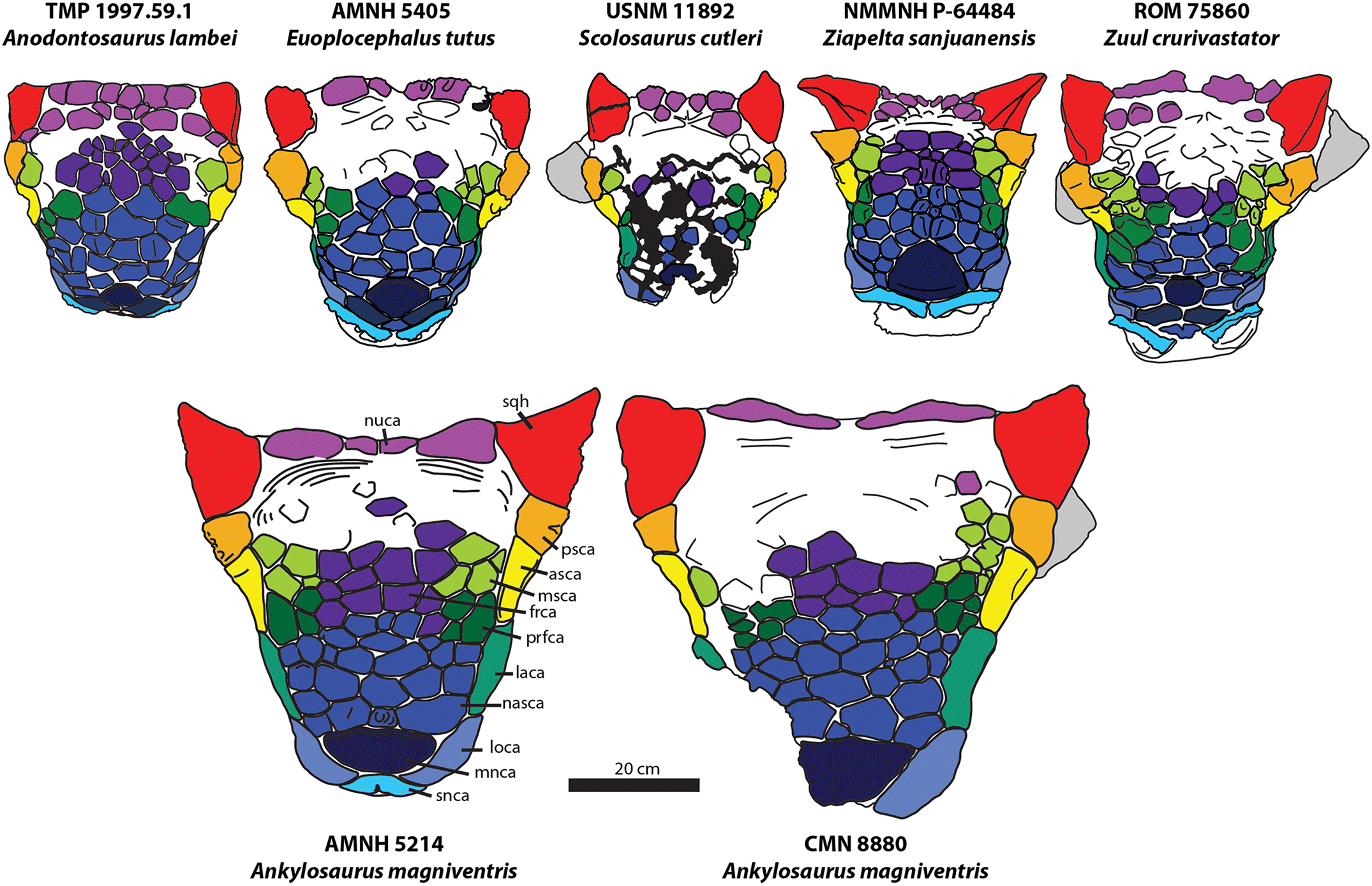
Ankylosaurus shares multiple cranial features in common with its close relatives Anodontosaurus, Dyoplosaurus, Euoplocephalus, Scolosaurus, Ziapelta, and Zuul (Vickaryous and Russell 2003; Arbour and Currie 2013a; Arbour et al. 2014a; Arbour and Evans 2017) (Figs. 2–4, Tables 1, 2, Supplementary Material 1). All of these taxa have (where preserved) cranial sculpturing characterized by rectangular to hexagonal frontonasal caputegulae, a large hexagonal median nasal caputegulum (except Ziapelta), a single loreal caputegulum, a single lacrimal caputegulum, and pyramidal squamosal and quadratojugal horns (Fig. 3). They are all similar in palatal view as well, with a broad premaxillary beak, deep but thin vomers, curved tooth rows, anterolaterally oriented pterygoid flanges, a short and robust braincase, and laterally oriented paroccipital processes (Fig. 2B). Ankylosaurus is easily distinguished from other derived Laramidian ankylosaurines (the ankylosaurins, Arbour and Currie 2016) based on aspects of its cranial ornamentation (Figs. 3, 4). Unlike Anodontosaurus, Euoplocephalus, and Scolosaurus, the keel on the anterior and posterior supraorbital osteoderms is continuous with the keel of the squamosal horn (Fig. 4). The squamosal horn is proportionately longer in Ankylosaurus compared with the squamosal horn of Euoplocephalus or Anodontosaurus, and is not posteroventrally curved as in Scolosaurus or lateroventrally curved as in Ziapelta (Fig. 3). It lacks the furrows present on the squamosal horn of Zuul. The median nasal caputegulum is proportionately large, similar to that in Euoplocephalus. In Ziapelta, the median nasal caputegulum is triangular with a posteriorly directed apex, and in Ankylosaurus the median nasal caputegulum is hexagonal, as in Anodontosaurus, Euoplocephalus, and Zuul. The loreal caputegulum in Ankylosaurus is proportionately larger than in other Laramidian ankylosaurins, expanding onto the anterior surface of the premaxillary beak and overhanging the lateral surface of the maxilla (Fig. 4).
Fig. 3.
Fig. 3. Skulls of Ankylosaurus compared with the ankylosaurins Anodontosaurus, Euoplocephalus, Scolosaurus, Ziapelta, and Zuul, in dorsal view, with cranial ornamentation colour coded for comparative purposes. asca, anterior supraorbital caputegulum; frca, frontal caputegulum; laca, lacrimal caputegulum; loca, loreal caputegulum; mnca, median nasal caputegulum; msca, middle supraorbital caputegulum; nasca, nasal caputegulum; nuca, nuchal caputegulum; psca, posterior supraorbital caputegulum; prfca, prefrontal caputegulum; snca, supranarial caputegulum; sqh, squamosal horn.
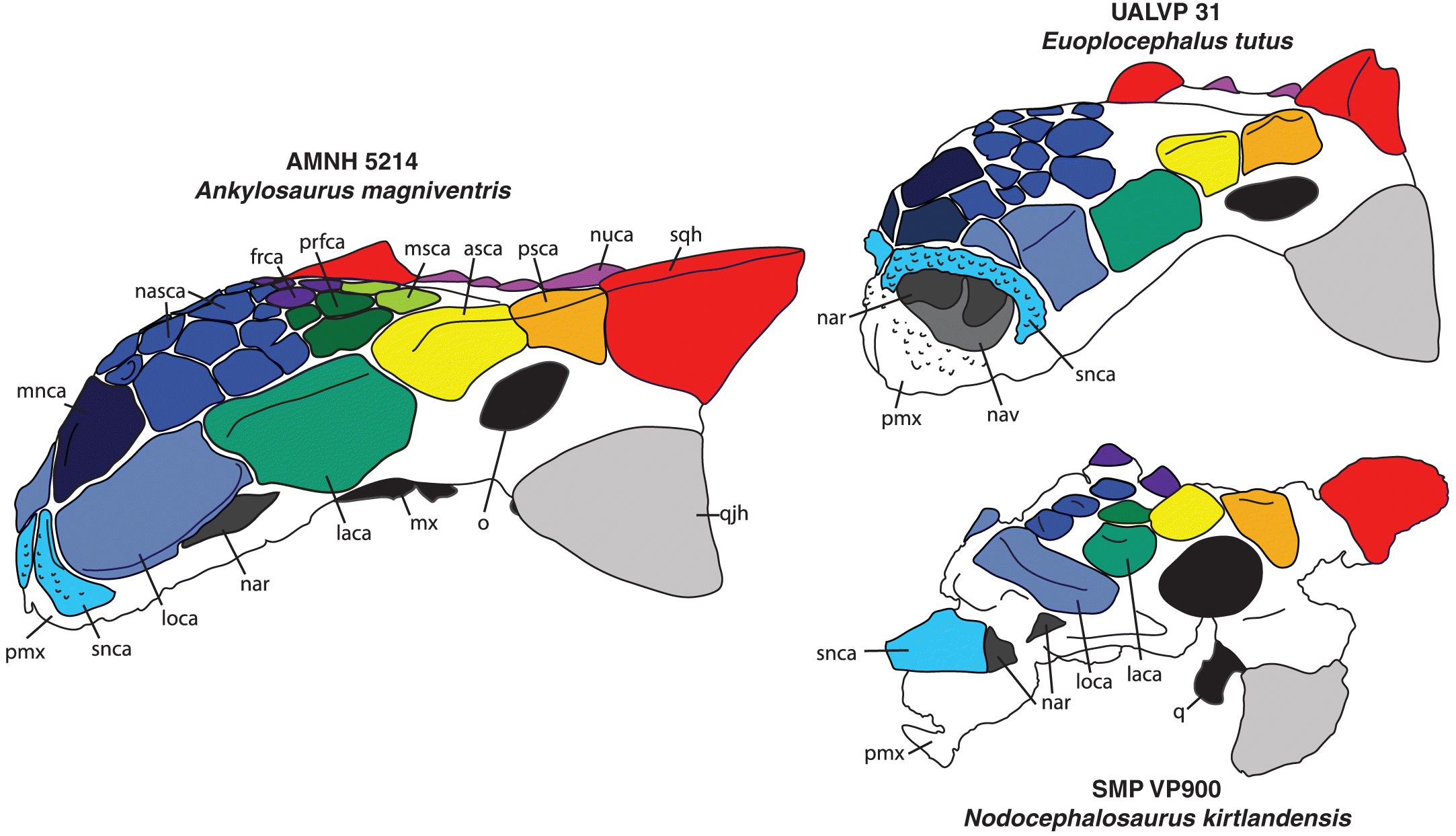
Fig. 4.
Fig. 4. Skulls of Ankylosaurus, Euoplocephalus, and Nodocephalosaurus, in left anterolateral view. In Euoplocephalus, the supranarial caputegulum is rugose and forms an arch over the anteriorly placed nasal vestibule. In Ankylosaurus, the nasal vestibule and external naris is roofed by the loreal caputegulum, and the supranarial caputegulum is small and flush with the premaxilla. The holotype of Nodocephalosaurus is poorly preserved in the premaxillary region, but the external nares are also posteriorly placed and may be partly roofed by a ridge-like loreal caputegulum; the supranarial caputegulum is also ridged and projects anteriorly. asca, anterior supraorbital caputegulum; frca, frontal caputegulum; laca, lacrimal caputegulum; loca, loreal caputegulum; mnca, median nasal caputegulum; msca, middle supraorbital caputegulum; mx, maxilla; nasca, nasal caputegulum; nar, external naris; nav, nasal vestibule; nuca, nuchal caputegulum; o, orbit; pmx, premaxilla; psca, posterior supraorbital caputegulum; q, quadrate; qjh, quadratojugal horn; snca, supranarial caputegulum; sqh, squamosal horn.
Table 1.
Table 1. Dimensions of ankylosaur tooth crown heights and skull lengths.
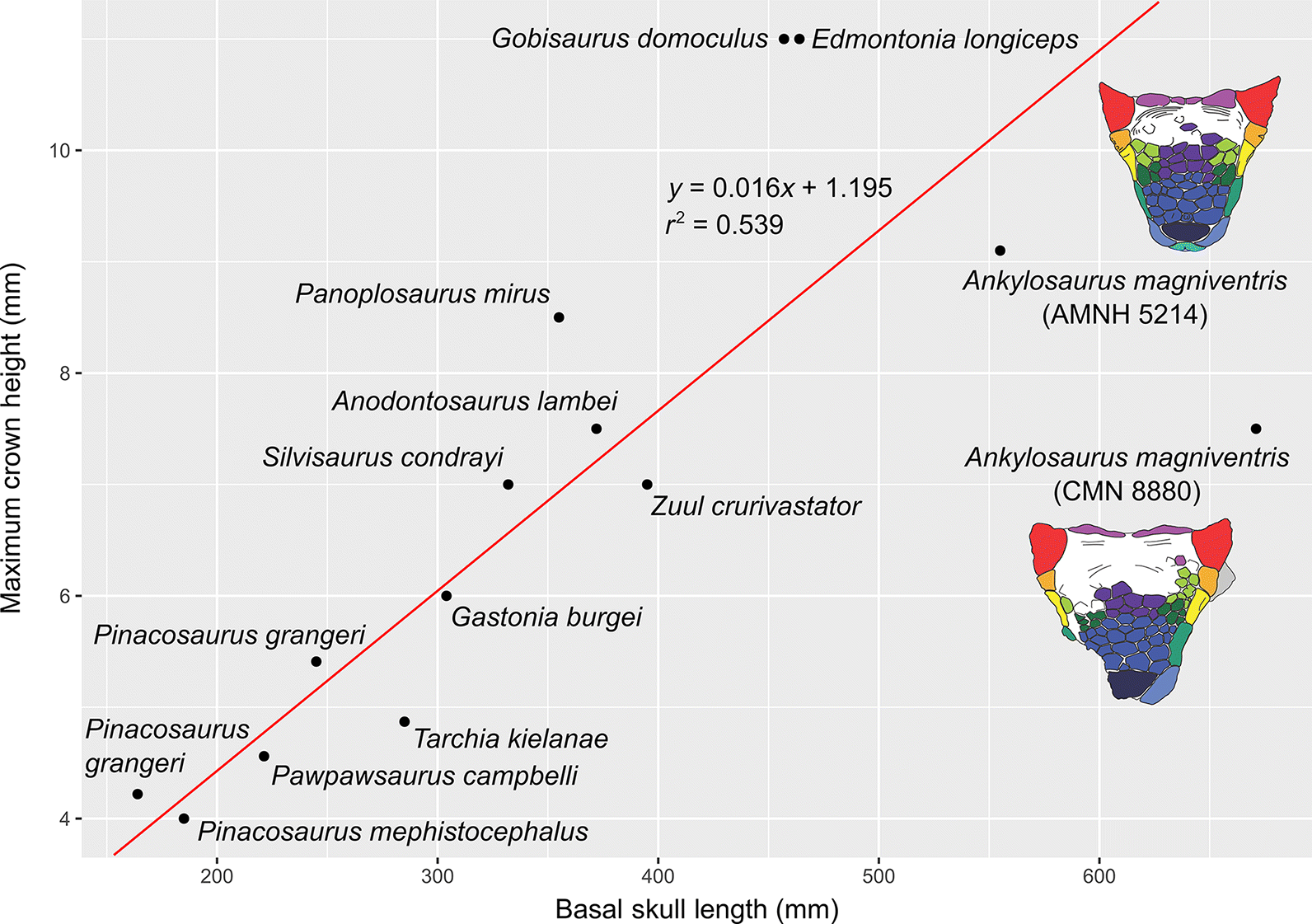
Carpenter (2004) provided a modern diagnosis for Ankylosaurus, noting multiple unique features of this taxon such as the lateral and posterior displacement of the external nares, and the greater number of maxillary and dentary teeth compared with other ankylosaurines. The greater number of teeth in Ankylosaurus does not appear to be simply a reflection of larger body size in Ankylosaurus; rather, a biplot of maximum (unworn) crown height vs. basal skull length in ankylosaurs illustrates that the teeth of Ankylosaurus are proportionally quite small (Table 1, Figs. 2G, 5), and that the jaws of the animal could therefore accommodate more of them. Tooth size appears to be highly variable in Ankylosaurus; the teeth of the largest skull (CMN 8880) are absolutely smaller than those of the smallest skull (AMNH 5214). The same variability is not seen in Pinacosaurus grangeri, the two representative specimens of which plot equidistant above the regression line (Fig. 5).
Some of the diagnostic characters Carpenter (2004) identified are also present in closely related ankylosaurins, which we comment on here. A posterodorsolaterally directed squamosal horn, posteroventrolaterally directed quadratojugal horn, and cranial ornamentation divided into polygons are present in all Laramidian ankylosaurins, not just Ankylosaurus (Fig. 3). In Ankylosaurus, the maximum width of the maxillary tooth rows at their posteriormost extent is similar to the maximum width of the premaxillary beak (Fig. 2B). This feature is also present in Anodontosaurus (e.g., TMP 1997.132.1), Ziapelta (Arbour et al. 2014a), and Zuul (Arbour and Evans 2017). Finally, the quadrate process of the pterygoid is directed posterolaterally in Ankylosaurus, and this feature is also present in Anodontosaurus, Euoplocephalus, and Zuul (Arbour and Currie 2013a; Arbour and Evans 2017).
The narial region in Ankylosaurus warrants extra attention, as it has undergone an extreme transformation relative to other Laramidian ankylosaurins. Ankylosaurines have unusual nasal anatomy, with complex nasal vestibules, convoluted looping nasal passages, and a high degree of vascularisation in the posterior portions of the airway (Hill et al. 2003; Witmer and Ridgely 2008; Miyashita et al. 2011). The border of the external naris in ankylosaurines is typically demarcated by a distinct edge on the premaxilla and the external surface of the supranarial caputegulum. Posterior to the external naris is the nasal vestibule, a concave region roofed by the nasals. The nasal vestibule contains one or more openings, the narial apertures, for the airway and sinuses (Hill et al. 2003). Some Mongolian taxa, such as Saichania and Pinacosaurus, possess three or more narial apertures (Hill et al. 2003; Arbour and Currie 2016); Anodontosaurus, Euoplocephalus, Zuul, and Ankylosaurus have a single folded narial aperture.
The external nares of most ankylosaurines are anteriorly to anterolaterally oriented (e.g., Maryańska 1977; Vickaryous et al. 2004; Arbour and Currie 2016), but the external nares in Ankylosaurus are located posteriorly relative to other ankylosaurs, and open ventrolaterally (Figs. 2, 4). Coombs (1971) suggested that this shift in external naris position resulted from the expansion of the nasals, Maryańska (1977) suggested it was overgrowth of cranial ornamentation, and Carpenter (2004) suggested it represented expansion of the internal sinuses. Unlike most other ankylosaurins, the external naris in Ankylosaurus is roofed by the loreal caputegulum, which takes on a bulbous, inflated appearance compared with that in Euoplocephalus, Anodontosaurus, Ziapelta, and Zuul (Figs. 2, 4). The supranarial caputegulae, which form an arched, rugose border above the external nares in most ankylosaurins, do not contribute to the border of the external nares at all in Ankylosaurus. Instead, they are reduced in size, and are flush with the skull rather than protruding anteriorly (Fig. 4). The loreal caputegulum covers part of the nasals in most ankylosaurins, and the supranarial caputegulae cover part of the premaxilla. As such, the interpretations presented by Coombs (1971) and Maryańska (1977) are both correct: the morphology of the caputegulae in this region suggests that the nasal bones, represented by the loreal caputegulae, have expanded anteriorly and pushed the supranarial caputegulae medially and anteriorly until they occupy only the tip of the snout.
Fig. 5.
Fig. 5. Reduced major axis biplot showing the relationship between ankylosaur tooth crown height (maxillary or dentary teeth) vs. basal skull length. Note that the teeth of Ankylosaurus are relatively small for its skull size. Basal skull length for CMN 8880 is as preserved and does not account for the broken premaxilla. Doing so would place the specimen even farther from the regression line.
One other ankylosaurine lacks anteriorly oriented external nares. Like Ankylosaurus, the external naris of Nodocephalosaurus is roofed by the loreal caputegulum, but this is ridge-like and laterally protruding, and the supranarial caputegulum is a prominent, anteriorly directed ridge (Sullivan 1999; Arbour and Currie 2016; Fig. 4). The boundaries of the external naris of Nodocephalosaurus are difficult to determine because of breakage in the only known specimen, but the naris likely faced only laterally, and not ventrolaterally. Nodocephalosaurus has not been recovered as a close relative of Ankylosaurus (e.g., Thompson et al. 2012; Arbour and Currie 2016), and so a posterior placement of the external nares was most likely independently evolved in these taxa.
The unusual placement of the external nares in Ankylosaurus is partially the result of anterior expansion of the loreal caputegulae (and therefore the nasals) and reduction of the premaxillae and the premaxillary contribution to the external naris. However, there also appears to have been a posterior shift in the external naris, and not just a lengthening of the snout anterior to the naris. The external naris in Ankylosaurus is bordered posteriorly by the lacrimal caputegulum, which covers the lacrimal and part of the maxilla in other ankylosaurins (Arbour and Currie 2013a). This caputegulum does not abut the external naris in Anodontosaurus, Euoplocephalus, Ziapelta, or Zuul. Additionally, in ventral view, the anterior edge of the tooth row (located entirely on the maxilla) is located at the midpoint of the external naris in Ankylosaurus but is aligned with the posterior corner of the supranarial caputegulae (and therefore external naris) in Euoplocephalus.
This expansion of the nasals and loreal caputegulae, and reduction of the dorsal surface of the premaxillae, may also have influenced the morphology of the premaxillary palate. Coombs (1971) noted that Ankylosaurus had a narrower beak than Euoplocephalus, which may in part reflect the widening of the nasals and loreal caputegulae as well as a narrowing of the premaxillary beak itself (Ősi et al. 2017). The expansion of the nasals and posterior shift of the external naris must also have modified the course of the internal nasal passages. AMNH 5895 is broken in such a way that the internal nasal passages are partially revealed in cross section. Witmer and Ridgely (2008) showed that the structures in Euoplocephalus previously identified by Coombs (1978a) as sinuses were instead a continuous looping airway; the channels visible in AMNH 5895 most likely also represent a continuous looping airway, although it is difficult to interpret the course of this airway at present in the absence of computed tomography data for this taxon.
All three of the known skulls of Ankylosaurus are very similar where the preserved areas overlap (Carpenter 2004; Fig. 2), and there is apparently slightly less intraspecific variation than that observed for Euoplocephalus or Anodontosaurus (Arbour and Currie 2013a), although the small sample size undoubtedly influences this. The only notable differences between AMNH 5214 and CMN 8880 are the overall size, the sharpness of the squamosal horn apex, and the distinctiveness of the nuchal caputegulae (Fig. 3). CMN 8880 has blunter squamosal horns than AMNH 5214, and the boundaries of the nuchal caputegulae are less distinct. Larger individuals of Euoplocephalus have more rounded squamosal horns than smaller individuals (Arbour and Currie 2013a), so this may represent ontogenetic variation in both Euoplocephalus and Ankylosaurus.
Postcranial anatomy
A unique trait of Ankylosaurus described by Carpenter (2004) concerns the morphology of the cervical half rings. Ankylosaurid half rings are unusual yoke-like structures that overlie the cervical and pectoral regions (Penkalski 2001; Arbour and Currie 2013a). There are usually two cervical half rings composed of an underlying band of non-osteodermal bone and an overlying set of six large osteoderms (Figs. 6A, 6B); numerous smaller osteoderms are sometimes present surrounding these major osteoderms. The cervical half ring band is made up of coossified, smooth, square, or rectangular segments with varyingly visible sutures in between the segments. Each segment of the band is topped by an osteoderm, and the distal segment on either side is usually enveloped along the distal edge by the osteoderm (Figs. 6A, 6B). Carpenter (2004) proposed that Ankylosaurus had “quarter” rings instead of half rings; i.e., there was a midline gap through the cervical half rings dividing them into two halves. We provide a reinterpretation of the preserved cervical half ring material in AMNH 5895 and argue that these represent more typical “half” rings, without a midline gap.
Fig. 6.
Fig. 6. Portions of Ankylosaurus (AMNH 5895) cervical half rings compared with the first cervical half ring of Euoplocephalus (AMNH 5406), and Ankylosaurus (AMNH 5895) postcervical osteoderms. (A) Complete (but distorted) first cervical half ring of Euoplocephalus (AMNH 5406) in ?anterior view, showing arrangement of osteoderms on the underlying band of bone, and (B) interpretive illustration. The medial and lateral osteoderm pairs sit atop flat band segments, but the distal osteoderms slightly envelop the tip of their band segments. (C) Portion of Ankylosaurus first cervical half ring with lateral and distal osteoderms in external view, and (D) interpretive illustration. (E) Portion of same half ring in internal view, and (F) interpretive illustration. Carpenter (2004) suggested that a second fragment of cervical half ring bearing a single osteoderm was a continuation of the segment figured here in C–F. In G–J, the cervical half ring fragment illustrated in C–F is located to the left, and the second fragment is located on the right. (G) These pieces (here in oblique anterior or posterior internal view) cannot be reunited along their broken edges and probably do not represent the same half ring; (H) interpretive illustration. (I) The osteoderm morphology of the fragment of cervical half ring with only a single preserved osteoderm (here in oblique anterior or posterior external view) is similar to that of the distal osteoderm of the more complete specimen, but is substantially larger, and thus probably represents a distal osteoderm from the second cervical half ring, which is typically larger in specimens of Euoplocephalus (Arbour and Currie 2013a); (J) interpretive illustration. (K) and (L) large postcervical osteoderms from Ankylosaurus (AMNH 5895), probably derived from medial positions along the back; the osteoderm in (L) is similar in shape to those immediately posterior to the medial cervical half ring osteoderms in Saichania (MPC 100/151) (Arbour and Currie 2013a, 2013b). b, cervical half ring band; dos-1, distal osteoderm of the first cervical half ring; dos-2, distal osteoderm of the second cervical half ring; los-1, lateral osteoderm of the first cervical half ring; mos-1, medial osteoderm of the first cervical half ring; p, plaster.
AMNH 5895 includes two large pieces of cervical half rings that do not join together in any orientation (Figs. 6C–6J). Comparison with cervical half rings from Anodontosaurus and Euoplocephalus (Penkalski 2001; Arbour and Currie 2013a; Figs. 6A, 6B) revealed that the larger cervical half ring fragment from AMNH 5895 includes the left lateral and distal osteoderms of the first cervical half ring (Figs. 6C–6F) and the smaller piece represents the left distal osteoderm from the second cervical half ring (Figs. 6I, 6J). Carpenter (2004) suggested that there was no evidence for an underlying bony band in Ankylosaurus, but the underlying band is visible on both fragments, although it is easier to discern on the fragment of the second cervical half ring. Overall, the cervical half rings of Ankylosaurus most closely resemble those of Euoplocephalus and Anodontosaurus (Arbour and Currie 2013a), which generally have osteoderms with oval bases and low to moderate keels; they are unlike those of Scolosaurus, which has circular osteoderms with low conical points (Arbour and Currie 2013a), or Ziapelta, which has rectangular osteoderms with tall keels (Arbour et al. 2014a). They differ even more from the cervical half rings of the more distantly related Shamosaurus, which has strongly anteriorly directed apices on the distal osteoderms (Arbour and Currie 2016).
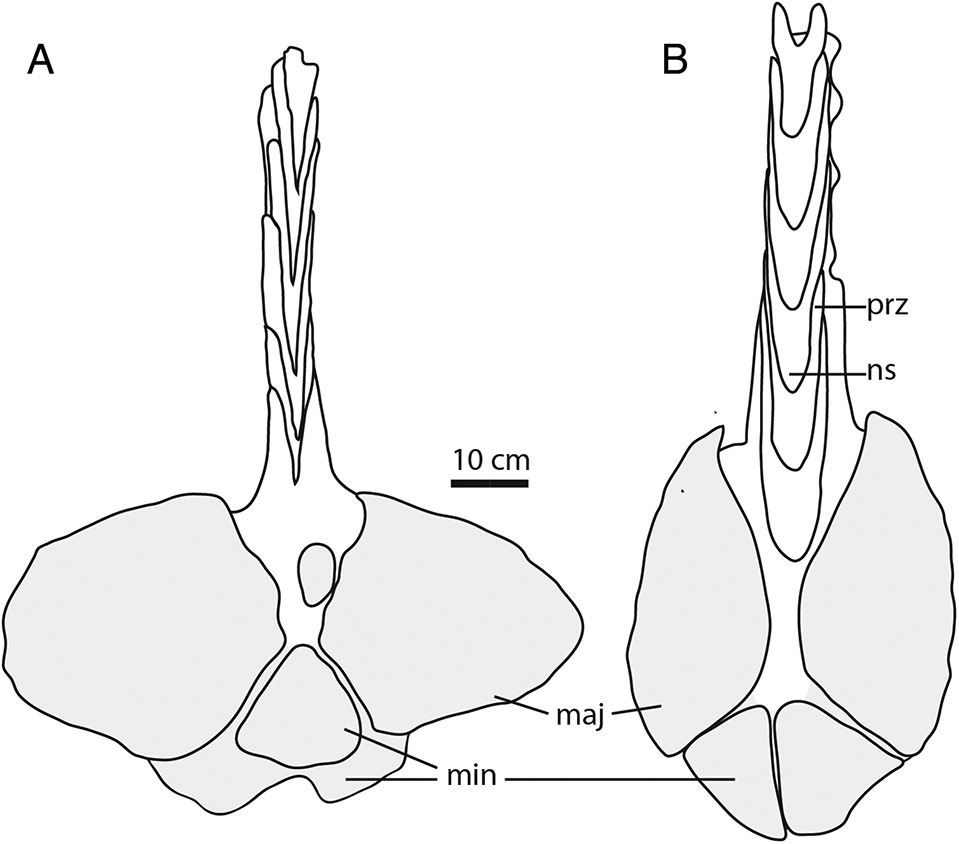
Although the tail of Ankylosaurus is poorly known, AMNH 5214 includes a portion of the tail club handle and a complete, well-preserved knob (Fig. 7). The handle vertebrae are twice as wide as those of Anodontosaurus and Dyoplosaurus, but are not longer. As such, the tail of Ankylosaurus may have been shorter relative to overall body length than in Anodontosaurus, Scolosaurus, Dyoplosaurus, or Zuul, or the tail may have had similar overall proportions but with a smaller tail club. The handle vertebrae of Ankylosaurus are unique among ankylosaurids, with U-shaped neural spines in dorsal view compared with the V-shaped neural spines in Anodontosaurus, Euoplocephalus, Pinacosaurus, Talarurus, and most other ankylosaurids (Maleev 1956; Maryańska 1969; Maryańska 1977; Tumanova 1987; Carpenter et al. 2011; Arbour and Currie 2013a; Loewen et al. 2013). This condition may simply reflect the wider handle of Ankylosaurus. Although the skull of AMNH 5214 is larger than skulls belonging to Anodontosaurus or Euoplocephalus, the tail club knob, at about 45 cm wide (measured from Carpenter 2004 using ImageJ), is not larger than the largest tail club knobs from Horseshoe Canyon Formation and Dinosaur Park Formation ankylosaurids (Anodontosaurus specimen AMNH 5425 is 59 cm wide, and an indeterminate tail club from Dinosaur Provincial Park, ROM 788, is 57 cm wide; Fig. 7). If CMN 8880, which is twice the width of the largest known Anodontosaurus skull, has a tail club knob twice the size of the largest Anodontosaurus tail club, then this specimen would have a knob width of 120 cm. This seems exceptionally large, and there must be an upward limit of knob mass that the handle vertebrae can support. Scaling the tail club knob of AMNH 5214 based on the skull width of CMN 8880 yields a knob width of about 57 cm, comparable to that of the largest Anodontosaurus and Euoplocephalus knobs.
Fig. 7.
Fig. 7. Partial tail clubs of (A) Anodontosaurus (AMNH 5245) and (B) Ankylosaurus (AMNH 5214) in dorsal view. maj, major knob osteoderm; min, minor knob osteoderm; ns, neural spine; prz, prezygapophysis.
Newly identified holotype material
Several trays of bone fragments are associated with AMNH 5895, including portions of the skull and cervical half rings that have not been identified or described in previous descriptions of Ankylosaurus. These include an otic capsule, maxilla fragment, right jugal, left jugal and quadratojugal, two sacral centra, and fragments of the cervical half rings (Fig. 8).
Fig. 8.
Fig. 8. Newly identified elements of AMNH 5895, holotype of Ankylosaurus magniventris. (A) Fragment of right maxilla, palatal view, anterior is to the right. (B) Left jugal and partial quadratojugal horn, lateral view. (C) Right jugal, lateral view. (D) Otic capsule, internal view. (E) Fragment of cervical half ring band with coossified osteoderm, anterior or posterior view. (F) Fragment of cervical half ring band segments with prominent suture, dorsal view. (G) Fragment of cervical half ring band, dorsal view. (H) Fragment of sacral rod, centra in ventral view. alv, alveolus; j, jugal; orb, lower border of orbit; os, osteoderm; qjh, quadratojugal horn; s, suture.
The maxilla fragment (Fig. 8A) is 6.3 cm long and 4.2 cm at its widest point. Seven alveoli are arranged diagonally along its long axis, but no teeth are preserved. Based on the angle of the alveoli and the robust projection at one end of the fragment, this might represent the posterior end of the right maxilla.
Parts of the lower portion of each orbit are present (Figs. 8B, 8C). A fragment representing the left jugal and partial quadratojugal horn (Fig. 8B) is 14.0 cm long and 13.1 cm high. The horn has a broad base, but the posterior edge is broken, as is the tip, making its overall shape unknown. As in other Ankylosaurus skulls, postocular caputegulae are absent along the quadratojugal horn. The presence of a thick wedge of plaster infill along the quadratojugal horn suggests that this portion may have been mounted with the rest of the skull at some point. A second fragment representing either the right jugal, right lacrimal, or portions of both (Fig. 8C) is 10.9 cm long and 5.0 cm high. It is transversely broad and shallowly concave.
An unusual fragment is robust, rounded, and includes a hemispherical concavity with a complex of large pores (Fig. 8D). This most likely represents one of the otic capsules, but because no other ankylosaur specimens are broken in such a way as to reveal this structure, it is difficult to compare with other species. Its greatest width is 5.2 cm and its greatest length is 6.4 cm. One side of the fragment completely lacks cortical bone surface and reveals a spongy texture. The opposite side includes a circular region approximately 1.3 by 2.3 cm, with four large foramina, and a central, pointed peak.
A poorly preserved fragment of the synsacrum (Fig. 8H) is 19.7 cm long, and only the ventral surface is preserved. One relatively complete centrum, and approximately one third of a second centrum, are preserved. They are elongate, slightly constricted at the midlength, and possess an indistinct, shallow midline furrow.
Multiple fragments of the cervical half rings (Figs. 8E–8G) were identified, although it is not possible to assign individual portions to either the first or second cervical half ring. These represent portions of the cervical half ring band segments which underlie the osteoderms. They have relatively smooth surfaces, with indistinct zigzag sutures marking the boundaries of band segments, and finger-like projections on the anterior and posterior edges.
Discussion
A new restoration of the osteoderm arrangement in Ankylosaurus
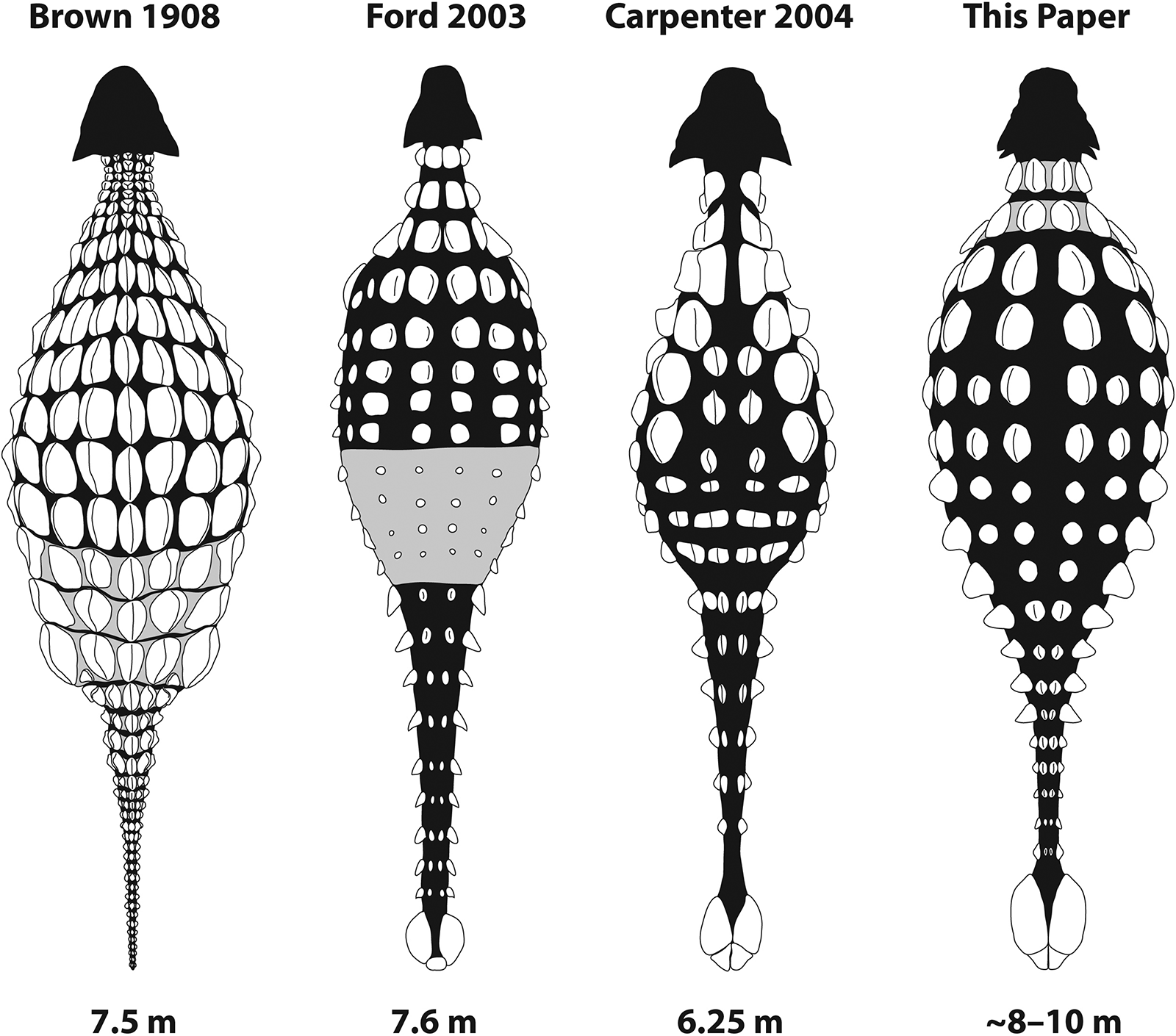
Brown (1908) included a restoration of the osteoderm arrangement in his original description of Ankylosaurus. Very few ankylosaur specimens were known at the time; Brown (1908) gave Ankylosaurus a suit of armour of closely packed thoracic osteoderms, interpreted the cervical half rings as originating from the pelvis and tail (partly inspired by the caudal rings in glyptodonts), and gave it a tapering, flexible tail lacking a tail club (Fig. 9). The 1910 discovery of AMNH 5214 provided the first evidence for a tail club in Ankylosaurus, and this structure was incorporated into later popular depictions of this ankylosaur, such as the 1947 Age of Reptiles mural by Rudolph Zallinger at the Yale Peabody Museum, or the 1964 World’s Fair Sinclair Dinoland sculpture. Popular depictions of Ankylosaurus also often conflated its armour with that of “Palaeoscincus” (now Edmontonia, a nodosaurid), giving it the large pectoral spikes known in the genus. Indeed, “Palaeoscincus” was also sometimes reconstructed with an Ankylosaurus-inspired tail club, including in a 1930 mural by Charles Knight at the Field Museum, most likely after the publication of an article showing two alternate interpretations of the tail of “Palaeoscincus” by Matthew (1922). Both Ford (2003) and Carpenter (2004) provided updated, modern restorations incorporating new knowledge about ankylosaur postcranial anatomy and osteoderm arrangements. Ford’s (2003) restoration shifted the caudal osteoderm rings to the cervical region (based on the presence of cervical half rings in many other ankylosaurids), included a tail club knob, increased the spacing between osteoderms in the thoracic and pelvic regions, varied the osteoderm size between the thoracic and pelvic regions, and hypothesized that a pelvic shield (coossified pelvic osteoderms) may have been present (Fig. 9). Carpenter (2004) similarly included cervical half rings and a tail club knob, and included a new arrangement of varying osteoderm shapes in the thoracic region (Fig. 9).
Fig. 9.
Fig. 9. Hypotheses of osteoderm morphologies and placements in Ankylosaurus magniventris. Brown (1908) proposed an arrangement in which the osteoderms are closely spaced and relatively uniform, with pelvic osteoderms united in coossified bands. Ford (2003) provided more space between the osteoderms overall, included a tail club knob, and hypothesized that a pelvic shield was present. Carpenter (2004) suggested that the cervical armour was arranged in quarter rings. We suggest that the cervical armour was united into more typical half rings, and provide an updated osteoderm arrangement based on preserved osteoderms in AMNH 5214, AMNH 5895, and AMNH 5866, and comparisons with related species.
We propose a new revision of the osteoderm arrangement in Ankylosaurus based on comparisons with its close relatives Anodontosaurus, Euoplocephalus, Dyoplosaurus, Scolosaurus, Zuul, and the more distant Mongolian relatives Saichania and Pinacosaurus (Fig. 9). Major changes in this restoration compared with those presented by Ford (2003) and Carpenter (2004) include revisions to the cervical half rings, the arrangement of the thoracic and pelvic osteoderms, and general body proportions. Our restoration is broadly congruent with that presented by Ford (2003).
Cervical half rings
All ankylosaurids that preserve cervical half ring material (e.g., Anodontosaurus (CMN 8530); Euoplocephalus (AMNH 5404, CMN 210, and UALVP 31); Pinacosaurus (IVPP V16854); Saichania (MPC 100/151); Scolosaurus (NHMUK R5161); Shamosaurus (PIN 3779/2); and Ziapelta (NMMNH P-64484)) demonstrate that there were at most two cervical half rings, and that these formed continuous U-shaped yokes over the dorsal part of the neck and anterior part of the shoulders. Ankylosaurus (AMNH 5895) had two large cervical half rings with six keeled, oval-based osteoderms on each half ring (Figs. 6, 9). The medial osteoderms are unknown in Ankylosaurus cervical half rings, but the lateral osteoderms are flat with a low keel.
Torso
The largest osteoderms (excluding smaller interstitial osteoderms and ossicles), like the cervical half rings, are arranged in transverse and longitudinal rows across most of the length of the body. Scolosaurus (NHMUK R5161) includes four or five transverse segments in the thoracic region delineated by creases in the skin impressions. Although no Ankylosaurus specimens preserve the integument in situ, Ankylosaurus likely had a similar osteoderm pattern consisting of transverse and longitudinal rows of osteoderms, with four or five transverse rows separated by skin creases.
The osteoderms immediately behind the second cervical half ring generally mimic the shape and arrangement of those in the half ring (e.g., MPC 100/151 and NHMUK R5161). In particular, a huge, roughly triangular-based osteoderm in AMNH 5895 (Fig. 6L) bears a strong similarity to the medial osteoderms in the first transverse row behind the second cervical half ring in MPC 100/151 (Arbour and Currie 2013b). Other large, generally oval osteoderms with low keels in AMNH 5895 and AMNH 5866 probably represent osteoderms in the dorsal thoracic region, although it is more difficult to place some of these precisely. In Scolosaurus (NHMUK R5161), dorsal osteoderms generally decrease in diameter posteriorly, so smaller flat osteoderms in AMNH 5866 were probably located more posteriorly.
MPC 100/1305, cf. Pinacosaurus, preserves in situ osteoderms on the left flank (Carpenter et al. 2011; Arbour and Currie 2013b). A typical flank osteoderm has a more square outline than those on the dorsal surface, and the keel is sigmoidal or kinked upward either anteriorly or posteriorly. Several osteoderms with this morphology are preserved in AMNH 5895 and AMNH 5866 (Carpenter 2004). MPC 100/1305 includes four longitudinal rows of flank osteoderms, and Ankylosaurus may have been similar.
Pelvic region
The osteoderms of the pelvic region are coossified into a continuous pelvic shield in the basal ankylosaurs Gastonia and Mymoorapelta, and in many nodosaurids (e.g., Taohelong, Nodosaurus, and Polacanthus), and are tightly appressed in the basal ankylosaurid Aletopelta (Lull 1921; Blows 1987; Kirkland and Carpenter 1994; Kirkland 1998; Yang et al. 2013; Arbour and Currie 2016). Pelvic osteoderms in ankylosaurines are preserved in several North American (Scolosaurus) and Asian (cf. Pinacosaurus and cf. Tarchia) specimens, and indicate that coossified pelvic shields were absent in this clade (Nopcsa 1928; Carpenter et al. 2011; Arbour et al. 2013). MPC 100/1305 has a mixture of small and large circular osteoderms on the pelvis, which, based on asymmetry in their arrangement on the skeleton, are probably somewhat displaced from their original position. NHMUK R5161 has three or four transverse rows of circular osteoderms that are relatively uniform in size and smaller than the osteoderms in the pectoral and thoracic regions. Osteoderms that can be confidently assigned to the pelvic region have not been identified in AMNH 5895, but Ankylosaurus may have possessed a similar suite of pelvic osteoderms as Scolosaurus based on their relatively close phylogenetic affinity.
Pelvic osteoderms in MPC 100/1305 are not present below the lower edge of the ilium. The lateral pelvic osteoderms, unlike the dorsal osteoderms, have a tall keel and take on a triangular profile in dorsal view. The largest of these osteoderms is located at the posterior corner of the ilium, and at least two more osteoderms of this shape, but decreasing in size, are present anteriorly. North American ankylosaurines do not appear to have had huge triangular lateral pelvic osteoderms, but both Dyoplosaurus and Scolosaurus preserve smaller triangular lateral pelvic osteoderms (Parks 1924; Nopcsa 1928; Arbour et al. 2009). A few osteoderms with this morphology are present in AMNH 5866 (Carpenter 2004), and these were probably located along the edges of the ilium.
Tail
In Asian and North American taxa where caudal osteoderms are preserved, such as cf. Pinacosaurus (PIN 614 and MPC 100/1305), Euoplocephalus (ROM 1930), Dyoplosaurus (ROM 784), and Zuul (ROM 75860), they are typically keeled or triangular and arranged in rows as on the rest of the body. Asian ankylosaurines preserve triangular lateral osteoderms along the tail club handle, but in North American specimens triangular osteoderms along the handle are so far only known in Zuul (Arbour and Evans 2017). AMNH 5895 and AMNH 5866 both include compressed triangular osteoderms (Carpenter 2004) that could have been located on the lateral sides of the pelvis or tail.
The tip of the tail in all ankylosaurines is enveloped by multiple osteoderms that obscure the terminal caudal vertebrae, forming the tail club knob (sensu Coombs 1995). AMNH 5214 is the only specimen of Ankylosaurus to preserve a tail club knob (Fig. 7). The knob is 60 cm long, 49 cm wide, and 19 cm high. It is semicircular in dorsal view, similar to the tail club knobs of Euoplocephalus and Scolosaurus, and unlike the pointed knob osteoderms of Anodontosaurus or the narrow and elongate knob of Dyoplosaurus.
Limbs
PIN 614 and MPC 100/1305 possess numerous ovoid, keeled, and sometimes teardrop-shaped osteoderms on the anterolateral sides of the upper and lower forelimbs. AMNH 5866 includes numerous small osteoderms with this morphology (Carpenter 2004), at least some of which were probably derived from the forelimbs. It is unclear if the hind limbs also bore osteoderms in ankylosaurines.
Size of Ankylosaurus
Like many other dinosaur taxa from the Hell Creek Formation, Ankylosaurus is substantially larger than its closest Campanian–Maastrichtian relatives (Russell and Manabe 2002; Longrich 2011; Lamanna et al. 2014; Brusatte and Carr 2016). Brown (1908) did not explicitly state any body size estimates for Ankylosaurus, but an illustration of the skeleton at 1/34 natural size was 22 cm long, and so Brown must have considered AMNH 5895 to be about 7.5 m in length. Carpenter (2004) proposed lengths of up to 6.25 m for CMN 8880 and up to 5.40 m for AMNH 5214 based on a review of the known skeletal material. Ford (2003) reconstructed Ankylosaurus with a length of approximately 7.6 m. We revisit these estimates with new data from more complete ankylosaurine skeletons here.
The skull of CMN 8880 is the largest ankylosaurid skull known: with a width across the supraorbitals of 66.2 cm, it is more than 70% larger than the largest known skulls of Anodontosaurus, Dyoplosaurus, Euoplocephalus, Scolosaurus, or Ziapelta (Table 2). AMNH 5895, a smaller individual, has a supraorbital width of 52.5 cm, making it about 35%–64% larger than the largest other ankylosaurins. The smallest individual, AMNH 5214, has a supraorbital width of 49.0 cm and premaxilla–occipital condyle width of 55.5 cm, and is therefore about 26%–50% larger than the largest skulls of other ankylosaurins.
No postcranial material is known for CMN 8880, but AMNH 5895 and AMNH 5214 preserve elements that can be compared with other ankylosaurins. AMNH 5337, a large Euoplocephalus, includes 10 free dorsal vertebrae (i.e., those not fused into the dorsosacral rod), as does AMNH 5895. Surprisingly, the dorsal vertebrae of AMNH 5895 are not noticeably larger than those of Euoplocephalus (Table 3). Only four caudal vertebrae are preserved in AMNH 5895 and their positions in the caudal sequence are unknown, but the largest of these is probably at least the fourth or fifth caudal vertebra based on the proportions of the transverse processes relative to the centrum and neural spine. The largest caudal vertebra in AMNH 5895 is much larger than any preserved in ROM 784 (Dyoplosaurus), and is similar in size to the largest caudals in ROM 1930 (Euoplocephalus). In the tail club handle, the vertebrae of AMNH 5214 are about the same length as those of AMNH 5245 (Anodontosaurus), ROM 784 (Dyoplosaurus), or ROM 788 (Euoplocephalus or Scolosaurus), but are about twice as wide. The femur of AMNH 5214 is between 12% and 25% longer than those of Dyoplosaurus, Euoplocephalus, and Scolosaurus, and the humerus is about 30% longer than UALVP 31 (Euoplocephalus) (Table 4).
Table 3.
Table 3. Dimensions of the cervical, dorsal, and caudal vertebrae of AMNH 5895, a mid-sized Ankylosaurus magniventris, compared with AMNH 5337, a large Euoplocephalus tutus.
Type
Position
AMNH 5895 Ankylosaurus magniventris
AMNH 5337 Euoplocephalus tutus
Centrum length (mm)
Centrum width (mm)
Centrum height (mm)
Centrum length (mm)
Centrum width (mm)
Centrum height (mm)
Cervical
2
61
95
82
—
—
—
Cervical
3
55
—
88
—
—
—
Cervical
4
68
—
93
—
—
—
Cervical
5
64
112
99
—
—
—
Cervical
6
73
135
109
—
—
—
Dorsal
1
105
98
102
104
113
103
Dorsal
2
123
—
—
117
108
106
Dorsal
3
127
113
98
120
109
105
Dorsal
4
132
116
107
125
109
112
Dorsal
5
134
105
108
121
116
110
Dorsal
6
127
120
114
115
116
107
Dorsal
7
126
121
111
112
120
112
Dorsal
8
120
120
110
110
106
108
Dorsal
9
121
125
111
108
124
113
Dorsal
10
120
131
114
113
128
107
Caudal
?
62
116
108
—
—
—
Caudal
?
66
113
109
—
—
—
Caudal
?
67
106
81
—
—
—
Caudal
?
62
100
81
—
—
—
Table 4.
Table 4. Comparative measurements and proportions of ankylosaurine postcranial elements.
Specimen
Width across supraorbitals (mm)
Scapula length (mm)
Humerus length (mm)
Femur length (mm)
Ankylosaurus magniventris
CMN 8880
662
—
—
—
AMNH 5895
525
615
—
—
AMNH 5214
490
—
542 / 536
670
Euoplocephalus tutus
ROM 1930
388
—
380
—
TMP 1979.14.164
385
—
—
—
AMNH 5405
372
—
—
—
AMNH 5337
365
—
415
—
AMNH 5404
—
—
403
535
UALVP 31
313
428
377
515
Anodontosaurus lambei
TMP 1997.132.1
355
—
399
—
Dyoplosaurus acutosquameus
ROM 784
355
—
—
562
Scolosaurus cutleri
NHMUK R5161
—
560
440
600
TMP 2001.42.1
263
—
—
430
Note: Measurements for NHMUK R5161 are from Nopcsa (1928).
In relatively complete ankylosaurs, such as PIN 614 and MPC 100/1305 (both cf. Pinacosaurus), ZPAL MgD I/113 (Ankylosaurinae indet.), ROM 784 (Dyoplosaurus), and ROM 75860 (Zuul), the tail club represents about 55%–64% of the total length of the tail (Table 5). In PIN 614 and MPC 100/1305, the tail represents about 49%–57% of the total length from the anterior of the cervical series to the distal end of the tail; both specimens lack skulls so our comparative lengths for Ankylosaurus are for the length of the body excluding the skull. AMNH 5214 preserves a portion of the tail club handle that is 1.21 m long, and in which seven vertebrae are present; ankylosaurins such as Dyoplosaurus have about 12 visible vertebrae in the tail club, so we can estimate that the complete tail club may have been about 2 m long, the tail may have been 3.12–3.63 m long, and the total body length (excluding the skull) could have been in the range of 5.47–7.40 m (with the skull included, 6.02–7.95 m). Given that the skull of CMN 8880 is 26% wider across the supraorbitals than AMNH 5214, and using the same proportion estimates as for AMNH 5214, we estimate that the length of CMN 8880 was between 6.89 and 9.32 m excluding the skull, and between 7.56 and 9.99 m including the skull (Table 5). We also attempted an estimate of the length of AMNH 5895 by drawing the lengths of the preserved elements to scale. Using measurements of the preserved skull and vertebrae of AMNH 5895 and the skull and tail club in AMNH 5214, estimating the length of the pelvis based on AMNH 5409 (a large pelvis from the Dinosaur Park Formation), and conservatively estimating the gaps between vertebrae and missing cervical and caudal vertebrae, we illustrated a length of approximately 6.5 m for AMNH 5895. Given that the vertebrae in AMNH 5895 do not differ substantially in size from other large ankylosaurin skeletons, a body length of nearly 10 m for a large Ankylosaurus is probably too long, but a length of up to 8 m is probably within reason.
Table 5.
Table 5. Comparative measurements and proportions of ankylosaurine skeletons.
Specimen
Tail length (cm)
Tail club length (cm)
Length from anterior end of cervical series to tail terminus (cm)
Skull width across supraorbitals (cm)
% of tail length represented by tail club
% of body length represented by tail
Estimated body length (excluding skull) (cm)
Estimated total body length (including skull) (cm)
The anterior portion of the tail of ROM 75860 is currently obscured by soft tissues, so total tail length may slightly exceed 300 cm.
b
AMNH 5245 is preserved in two sections: the knob and a substantial portion of the handle measuring 88 cm long, and an additional fragment of the handle measuring 43 cm long; the anterior edge is not preserved on the fragment and so the total length of the tail club may have been even greater.
Using similar proportion estimates for other relatively complete ankylosaurines indicates that Ankylosaurus was the largest ankylosaurine (and possibly even the largest ankylosaur), but a few other species approach our lowest length estimates for Ankylosaurus. PIN 614 and MPC 100/1305 (both cf. Pinacosaurus) have neck-to-tail lengths of 3.27 and 3.66 m, respectively, and we estimate a length of between 3.58 and 4.16 m for ROM 784 (Dyoplosaurus). Based on a relatively complete tail club (AMNH 5245), we estimate that Anodontosaurus reached at least 4 m in length. The holotype of Scolosaurus (NHMUK R5161) lacks the skull and tail club but still measures nearly 4 m in length (Nopcsa 1928), and most likely approached 6 m in total. No specimens of Euoplocephalus are complete enough to make a confident length estimate (Arbour and Currie 2013a), but based on comparisons with skull and limb proportions for Anodontosaurus and Scolosaurus, a large Euoplocephalus may have been 5–6 m long. A large indeterminate ankylosaurine taxon from Mongolia (ZPAL MgD I/113) has a 3.10 m long tail and may also have exceeded 6 m in total body length. Finally, Zuul is estimated to have been about 6 m long, based on the length of the nearly complete, partially articulated skeleton in the field (Arbour and Evans 2017). Large individuals of Ankylosaurus almost certainly exceeded 6 m in length.
To calculate the mass of Ankylosaurus, we used the QE function in the MASSTIMATE v. 1.3 package of Campione (2016) in R version 3.2.3 (R Core Team 2015). This package estimates body mass from combined stylopodial shaft circumferences using a scaling equation described by Campione and Evans (2012). For AMNH 5214, the smallest known Ankylosaurus (humeral shaft circumference = 315 mm, femoral shaft circumference = 363 mm), the estimated mass is 4.78 ± 1.22 t. For comparison, the estimated mass of AMNH 5404, a large specimen of Euoplocephalus (humeral shaft circumference = 244 mm, femoral shaft circumference = 278 mm), is 2.33 ± 0.60 t. Thus, Ankylosaurus appears to have been a substantially bulkier animal, even if its total body length did not greatly exceed those of its closest relatives (which is consistent with standard allometric scaling relationships of the vertebrate skeleton).
How heavy was the largest known Ankylosaurus, CMN 8880? With only an isolated skull available, it is difficult to be certain. However, it is possible to hazard a first approximation by isometrically scaling up stylopodial shaft circumferences from AMNH 5214 using width between the supraorbitals as a common reference. (Stylopodial shaft circumference very likely scaled allometrically in the genus, but there are too few specimens to determine a scaling coefficient.) In this way, we estimate that CMN 8880 might have weighed approximately 7.95 ± 2.04 t—about as massive as a large male African elephant (Loxodonta africana) (Colbert 1993)—but these values must obviously be regarded with due scepticism. Other published mass estimates for Ankylosaurus (Paul 1997), presumably using the smaller and more complete AMNH 5214 as reference, place the animal at approximately 6 t, although methodological details in these studies are lacking. Seebacher (2001) estimated a mass of just ∼1.7 t using his polynomial technique, which strikes us as excessively small for an animal of otherwise elephantine proportions.
Palaeoecology of Ankylosaurus
Ankylosaurus was a rare component of late Maastrichtian fossil assemblages: Lyson and Longrich (2011) found it made up <0.05% of associated dinosaur specimens from the Hell Creek, Lance, and equivalent formations, Wilson (2008) found it was less than 0.05% of isolated specimens from microvertebrate localities, and Horner et al. (2011) did not recover any specimens from the middle or upper third of the Hell Creek Formation and found it was 5% of the fauna in the lower third. Ankylosaurus may have been ecologically rare, or it may have been an infrequent visitor to the coastal plain depositional environments where it was more likely to fossilize (similar comments were made by Mallon and Anderson (2014b) concerning ankylosaurids from the older Dinosaur Park Formation). Carpenter (2004) suggested that the late Maastrichtian Edmontonia sp. was ecologically separated from Ankylosaurus, based on the fact that the former is more abundant in coastal “lowland” settings; however, no data were provided in support of this assertion. Where the two genera overlap in the lower Hell Creek Formation and elsewhere (Carpenter and Breithaupt 1986; Horner et al. 2011), it is likely that they practiced some form of dietary niche partitioning, as revealed by differences in beak and tooth morphology (Carpenter 1982b, 2004; Mallon and Anderson 2014a, 2014b).
Although rarer than other herbivorous dinosaurs of Lancian age, the immense size of Ankylosaurus would have made it a formidable herbivore. Its low build and wide beak would have enabled it to ingest abundant, low-growing plant matter (Mallon and Anderson 2014a; Ősi et al. 2017)—possibly ferns and low-growing shrubs, which were readily available in the Late Cretaceous (Wing et al. 1993). Hummel et al. (2008) estimated that the mean amount of metabolizable energy (ME) available from ferns is 7.7 MJ/kg of dry matter. Assuming that a large, ectothermic individual of Ankylosaurus required 55 kJ ME/kg BM0.75 per day (Hummel et al. 2008), this would work out to ∼6 kg of dry ferns per day (∼2 t per year). Hummel et al. (2008) considered such paltry requirements for such a large herbivore unrealistic. An endothermic individual, requiring 10× the amount of energy, would have eaten ∼60 kg per day (∼20 t per year). This is approximately equivalent to that predicted for a large elephant subsisting on dry vegetation (Laws 1970). Of course, these nutritional requirements could have been attained more efficiently through the added consumption of energy-rich fruiting bodies, for which the miniscule, cusp-like teeth (Mallon and Anderson 2014b), and comparably selective beak shape of Ankylosaurus (particularly in relation to Euoplocephalus; Ősi et al. 2017) were apparently well adapted. Small and nutritious invertebrates may also have supplemented the diet on occasion (see below). Undoubtedly, given the comparatively little chewing that Ankylosaurus would have performed (Colbert 1993; Ősi et al. 2017), foraging time would have taken up a less substantial portion of the day than for an elephant; food processing would have occurred primarily via fermentation in the expanded gut (Bakker 1986; Farlow 1987).
Given the propensity of elephants to congregate, fell trees, and strip tree bark, they are commonly considered allogenic ecosystem engineers, capable of creating open habitats through their otherwise destructive foraging activities (Jones et al. 1994). This phenomenon has been shown to influence the structure of smaller ungulate guilds (Fritz et al. 2002). In light of their comparable size and (arguably) energetic requirements, were Ankylosaurus ecosystem engineers in the same way? We suggest not. Although some ankylosaurs are known to have congregated at a young age (Burns et al. 2011), the same does not appear to be true of the larger adults, as partial skeletons collected from western North America are almost always found as isolated individuals, and not as bone beds or associations of individuals. Ankylosaurs also did not possess an elevated centre of mass as elephants do to aid in systematic tree felling (Mallon et al. 2013). Lastly, ankylosaurs were certainly not capable of chewing tough tree bark in the same way as elephants (Mallon and Anderson 2014b), and so were unlikely to practice bark stripping and the indirect destruction of trees via pathogenic infection. For these reasons, although remarkable herbivores, we do not think ankylosaurs had the same modifying influence on their respective ecosystems. The more abundant and capable hadrosaurids were most likely to be the ecosystem engineers of their day.
Although ankylosaurines are typically categorized as herbivores, the unusual narial anatomy of Ankylosaurus could reflect a change in diet or feeding strategy relative to other ankylosaurs and warrants further discussion. Posterior retraction of the external nares is a well-documented phenomenon in aquatic vertebrates such as cetaceans (Berta et al. 2014) and metriorhynchid crocodyliformes (Young et al. 2010), and retraction of the external naris is also associated with the development of a proboscis or enlarged nasal cartilages, such as in proboscideans, tapirs, saiga antelope (Saiga), and moose (Alces) (Witmer et al. 1999; Clifford and Witmer 2004a, 2004b). Neither of these seem to be likely functional interpretations of the morphology in Ankylosaurus, especially because retraction of the naris for aquatic- or proboscis-bearing species is usually achieved via reduction, not expansion, of the nasals, or through expansion of the premaxilla, and Ankylosaurus lacks any obvious aquatic adaptations otherwise. In addition to the posterior shift in external naris position, the ventrolateral orientation and wide separation of the external nares of Ankylosaurus are highly unusual among amniotes. Clifford and Witmer (2004a) hypothesized that the widely separated nostrils of moose (Alces) may allow for stereolfaction. This ability is poorly understood in amniotes and was probably not a major selective pressure for the observed narial anatomy in moose (Clifford and Witmer 2004a), and while stereolfaction may have been possible in Ankylosaurus, the data are too scant at present to infer this as the underlying cause of the changes in narial anatomy in this genus.
An alternate explanation for the smaller, posteriorly set, and dorsally roofed external nares in Ankylosaurus is that this feature evolved as a response to subterranean rooting. Extant mammals that engage in rooting or grubbing for food, such as suids, aardvarks, and armadillos, tend to have long snouts tipped by a flat disc with forward-facing nostrils, which is unlike the morphology observed in Ankylosaurus. However, ventrally placed or dorsally roofed external nares, combined with a solid, blunt, shovel-like rostrum, are present in several fossorial squamates of the clades Amphisbaenia and Scolecophidia (e.g., Diplometopon zarudnyi, Liotyphlops albirostris, Rhineura floridana, Typhlops jamaicensis; Maisano et al. 2006; Rieppel et al. 2009; Gauthier et al. 2012). While we do not suggest that Ankylosaurus was fossorial, it is possible that the similarities in rostral anatomy between this ankylosaur, amphisbaenians, and scolecophidians may reflect convergent adaptations for earth moving behaviours. Combined with other unusual cranial adaptations such as deep vomers that subdivide the oral cavity, an inferred robust, muscular tongue based on the presence of large, plate-like hyoglossia in Saichania and Pinacosaurus (Maryańska 1977; Hill et al. 2015), and relatively low tooth formation rates compared with other ornithischians (Hill et al. 2015), as well as the recent discovery of a skeleton of Liaoningosaurus with fish preserved in its thoracic cavity (Ji et al. 2016), an omnivorous or at least not strictly folivorous diet seems increasingly plausible for some ankylosaurids. Alternatively (or additionally), Ankylosaurus may have foraged for roots and tubers. The propensity for some ankylosaurs to dig was also suggested by Coombs (1978b) based on the inferred muscle action of the forelimb and shoulder joint morphology. This behaviour is also consistent with the presence of a syncervical in some species (VanBuren and Evans 2017), although the condition is unknown in Ankylosaurus.
Conclusions
Despite being the namesake of its entire suborder, Ankylosaurus was far from a representative ankylosaur. It was, in many respects, quite bizarre, having modified the dentition and narial region beyond convention, and having attained a much larger body size than its predecessors. Perhaps the small teeth were adapted for dealing with small fruiting bodies or invertebrates. Perhaps the lateral displacement of the narial passages signals an evolutionary shift in the function of the internal nasal passages, in olfactory ability, or in diet. Although it is possible to speculate about the adaptiveness of these features, choosing among the alternatives is presently impossible. In view of the fact that Ankylosaurus remains among the more poorly represented ankylosaurs, further questions concerning its anatomy and functional morphology abound. Why are the teeth so variable in size? What functional constraints limited the size of the tail club? These and other questions merit consideration going forward.
Institutional abbreviations
AMNH, American Museum of Natural History, New York, New York, USA; CCM, Carter County Museum, Ekalaka, Montana; CEUM, Utah State University Eastern Prehistoric Museum, Price, Utah, USA; CMN, Canadian Museum of Nature, Ottawa, Ontario, Canada; FWMSH, Fort Worth Museum of Science and History, Fort Worth, Texas, USA; IMM, Inner Mongolia Museum, Hohhot, Nei Mongol, China; INBR, Victor Valley Museum, Apple Valley, California, USA; IVPP, Institute for Vertebrate Paleontology and Paleoanthropology, Beijing, China; MPC, Paleontological Center, Mongolian Academy of Sciences, Ulaanbaatar, Mongolia; NHMUK, Natural History Museum, London, United Kingdom; NMMNH, New Mexico Museum of Natural History and Science, Albuquerque, New Mexico, USA; PIN, Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia; ROM, Royal Ontario Museum, Toronto, Ontario, Canada; SMP, State Museum of Pennsylvania, Harrisburg, Pennsylvania, USA; SMU, Shuler Museum of Paleontology at Southern Methodist University, Dallas, Texas, USA; TMP, Royal Tyrrell Museum of Paleontology, Drumheller, Alberta, Canada; UALVP, University of Alberta Laboratory for Vertebrate Paleontology, Edmonton, Alberta, Canada; UCMP, University of California Museum of Paleontology, Berkeley, California, USA; UW, University of Wyoming Geological Museum, Laramie, Wyoming, USA; UKMNH, University of Kansas Museum of Natural History, Lawrence, Kansas, USA; ZPAL, Zaklad Paleobiologii, Polish Academy of Sciences, Warsaw, Poland.
Acknowledgements
A. McDonald and S. Swan helped to conserve skull of CMN 8880. A. McDonald, M. Livius, and S. en Pan created the 3D model of CMN 8880, with additional input from R. McCrea. Collections access was graciously provided by M. Norell and C. Mehling (AMNH), J. Bartlett and R. Barrick (CEUM), L. Ballinger (FWMSH), Xu X. and Zheng F. (IVPP), K. Tsogtbaatar and Chinzorig Tsogtbaatar (MPC), S. Chapman (NHMUK), S. Lucas (NMMNH), T. Tumanova (PIN), R. Sullivan (SMP), B. Strilisky (TMP), and M. Borsuk-Białynicka (ZPAL). J. Baardse, T. Parker, C. Masnaghetti, and N. Turinetti (Urvogel Games LLC) created a 3D digital life restoration of Ankylosaurus in conjunction with VMA, which served as the basis for the illustration in Fig. 9. Thanks to P. Currie, D. Evans, M. Burns, J. Farlow, and C. Mehling for discussions, and to S. Maidment, an anonymous reviewer, and editor C. Clemente for comments and suggestions that improved this manuscript. Funding for this research was provided by the Natural Sciences and Engineering Research Council, an Alberta Ingenuity Studentship, the Dinosaur Research Institute, and Urvogel Games LLC (to VMA).
References
Arbour VM, and Currie PJ. 2013a. Euoplocephalus tutus and the diversity of ankylosaurid dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA. PLoS ONE, 8: e62421.
Arbour VM, and Currie PJ. 2013b. The taxonomic identity of a nearly complete ankylosaurid dinosaur skeleton from the Gobi Desert of Mongolia. Cretaceous Research, 46: 24–30.
Arbour VM, and Currie PJ. 2016. Systematics, phylogeny and palaeobiogeography of the ankylosaurid dinosaurs. Journal of Systematic Palaeontology, 14: 385–444.
Arbour VM, and Evans DC. 2017. A new ankylosaurine dinosaur from the Judith River Formation of Montana, USA, based on an exceptional skeleton with soft tissue preservation. Royal Society Open Science, 4: 161086.
Arbour VM, Burns ME, and Sissons RL. 2009. A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus. Journal of Vertebrate Paleontology, 29: 1117–1135.
Arbour VM, Lech-Hernes NL, Guldberg TE, Hurum JH, and Currie PJ. 2013. An ankylosaurid dinosaur from Mongolia with in situ armour and keratinous scale impressions. Acta Palaeontologica Polonica, 58: 55–64.
Arbour VM, Burns ME, Sullivan RM, Lucas SG, Cantrell AK, Fry J, et al. 2014a. A new ankylosaurid dinosaur from the Upper Cretaceous (Kirtlandian) of New Mexico with implications for ankylosaurid diversity in the Upper Cretaceous of western North America. PLoS ONE, 9: e108804.
Arbour VM, Currie PJ, and Badamgarav D. 2014b. The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia. Zoological Journal of the Linnean Society, 172: 631–652.
Blows WT. 2001. Dermal armor of the polacanthine dinosaurs. In The armored dinosaurs. Edited by K Carpenter. Indiana University Press, Bloomington, Indiana. pp. 363–385.
Brown B. 1908. The Ankylosauridae, a new family of armored dinosaurs from the Upper Cretaceous. Bulletin of the American Museum of Natural History, 24: 187–201.
Burns ME. 2009. An armoured dinosaur of the Frenchman Formation: using osteoderms to study ankylosaur geographic and stratigraphic distribution. In Frenchman Formation Terrestrial Ecosystem Conference, Royal Saskatchewan Museum Contribution to Science Number 12. pp. 11–13.
Burns ME, Currie PJ, Sissons RL, and Arbour VM. 2011. Juvenile specimens of Pinacosaurus grangeri Gilmore, 1933 (Ornithischia: Ankylosauria) from the Late Cretaceous of China, with comments on the specific taxonomy of Pinacosaurus. Cretaceous Research, 32: 174–186.
Campione NE. 2016. MASSTIMATE: body mass estimation equations for vertebrates. R package version 1.3 [online]: Available from CRAN.R-project.org/package=MASSTIMATE.
Campione NE, and Evans DC. 2012. A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods. BMC Biology, 10: 60.
Carpenter K. 1982a. Baby dinosaurs from the Late Cretaceous Lance and Hell Creek formations and a description of a new species of theropod. Contributions to Geology, University of Wyoming, 20: 123–134.
Carpenter K. 1982b. Skeletal and dermal armor reconstruction of Euoplocephalus tutus (Ornithischia: Ankylosauridae) from the Late Cretaceous Oldman Formation of Alberta. Canadian Journal of Earth Sciences, 19: 689–697.
Carpenter K. 2004. Redescription of Ankylosaurus magniventris Brown 1908 (Ankylosauridae) from the Upper Cretaceous of the Western Interior of North America. Canadian Journal of Earth Sciences, 41: 961–986.
Carpenter K, and Breithaupt B. 1986. Latest Cretaceous occurrence of nodosaurid ankylosaurs (Dinosauria, Ornithischia) in western North America and the gradual extinction of the dinosaurs. Journal of Vertebrate Paleontology, 6: 251–257.
Carpenter K, Hayashi S, Kobayashi Y, Maryańska T, Barsbold R, Sato K, et al. 2011. Saichania chulsanensis (Ornithischia, Ankylosauridae) from the Upper Cretaceous of Mongolia. Palaeontographica Abteilung A, 294: 1–61.
Cignoni P, Callieri M, Corsini M, Dellepiane M, Ganovelli F, and Ranzuglia G. 2008. MeshLab: an open-source mesh processing tool. In Sixth Eurographics Italian Chapter Conference, Salerno, Italy, 2–4 July 2008. pp. 129–136.
Clifford AB, and Witmer LM. 2004a. Case studies in novel narial anatomy: 2. The enigmantic nose of moose (Artiodactyla: Cervidae: Alces alces). Journal of Zoology, 262: 339–360.
Clifford AB, and Witmer LM. 2004b. Case studies in novel narial anatomy: 3. Structure and function of the nasal cavity of saiga (Artiodactyla: Bovidae: Saiga tatarica). Journal of Zoology, 264: 217–230.
Coombs WP Jr. 1990. Teeth and taxonomy in ankylosaurs. In Dinosaur systematics: approaches and perspectives. Edited by K Carpenter and PJ Currie. Cambridge University Press, Cambridge, UK. pp. 269–279.
Coombs WP Jr. 1995. Ankylosaurian tail clubs of middle Campanian to early Maastrichtian age from western North America, with description of a tiny club from Alberta and discussion of tail orientation and tail club function. Canadian Journal of Earth Sciences, 32: 902–912.
Dixon JR. 2000. Amphibians and reptiles of Texas: with keys, taxonomic synopses, bibliography, and distribution maps. 2nd edition. Texas A&M University Press, College Station, Texas. 425 p.
Eaton TH Jr. 1960. A new armored dinosaur from the Cretaceous of Kansas. The University of Kansas Paleontological Contributions: Vertebrata 8. The University of Kansas, Lawrence, Kansas. pp. 1–24.
Ford TL. 2003. A new look at the armor of Ankylosaurus, just how did it look? In The Mesozoic in Wyoming. Edited by DE Brown. The Geological Museum, Casper College, Casper, Wyoming. pp. 48–68.
Fritz H, Duncan P, Gordon IJ, and Illius AW. 2002. Megaherbivores influence trophic guilds structure in African ungulate communities. Oecologia, 131: 620–625.
Gauthier JA, Kearney M, Maisano JA, Rieppel O, and Behlke ADB. 2012. Assembling the squamate tree of life: perspectives from the phenotype and the fossil record. Bulletin of the Peabody Museum of Natural History, 53: 3–308.
Godefroit P, Pereda Suberbiola X, Li H, and Dong ZM. 1999. A new species of the ankylosaurid dinosaur Pinacosaurus from the Late Cretaceous of Inner Mongolia (P.R. China). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 69(Suppl.): 17–36.
Hill RV, Witmer LW, and Norell MA. 2003. A new specimen of Pinacosaurus grangeri (Dinosauria: Ornithischia) from the Late Cretaceous of Mongolia: ontogeny and phylogeny of ankylosaurs. American Museum Novitates, 3395: 1–29.
Hill RV, D’Emic MD, Bever GS, and Norell MA. 2015. A complex hyobranchial apparatus in a Cretaceous dinosaur and the antiquity of avian paraglossalia. Zoological Journal of the Linnean Society, 175: 892–909.
Horner JR, Goodwin MB, and Myhrvold N. 2011. Dinosaur census reveals abundant Tyrannosaurus and rare ontogenetic stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PLoS ONE 6: e16574.
Hummel J, Gee CT, Südekum KH, Sander PM, Nogge G, and Clauss M. 2008. In vitro digestibility of fern and gymnosperm foliage: implications for sauropod feeding ecology and diet selection. Proceedings of the Royal Society of London B: Biological Sciences, 275: 1015–1021.
Ji Q, Wu X, Cheng Y, Ten F, Wang X, and Ji Y. 2016. Fish hunting ankylosaurs (Dinosauria, Ornithischia) from the Cretaceous of China. Journal of Geology, 40: 183–190.
Kirkland JI. 1998. A polacanthine ankylosaur (Ornithischia: Dinosauria) from the Early Cretaceous (Barremian) of eastern Utah. New Mexico Museum of Natural History and Science Bulletin, 14: 271–281.
Kirkland JI, and Carpenter K. 1994. North America’s first pre-Cretaceous ankylosaur (Dinosauria) from the Upper Jurassic Morrison Formation of western Colorado. BYU Geology Studies, 40: 25–42.
Lamanna MC, Sues H-D, Schachner ER, and Lyson TR. 2014. A new large-bodied oviraptorosaurian theropod dinosaur from the latest Cretaceous of western North America. PLoS ONE, 9: e92022.
Leahey LG, Molnar RE, Carpenter K, Witmer LM, and Salisbury SW. 2015. Cranial osteology of the ankylosaurian dinosaur formerly known as Minmi sp. (Ornithischia: Thyreophora) from the Lower Cretaceous Allaru Mudstone of Richmond, Queensland, Australia. PeerJ, 3: e1475.
Lillegraven JA, and Eberle JJ. 1999. Vertebrate faunal changes through Lancian and Puercan time in southern Wyoming. Journal of Paleontology, 73: 691–710.
Loewen MA, Burns ME, Getty MA, Kirkland JI, and Vickaryous MK. 2013. A review of the Late Cretaceous ankylosaurian dinosaurs from the Grand Staircase of southern Utah. In The Late Cretaceous in Utah. Edited by AL Titus. Indiana University Press, Bloomington, Indiana. pp. 445–462.
Lyson TR, and Longrich NR. 2011. Spatial niche partitioning in dinosaurs from the latest Cretaceous (Maastrichtian) of North America. Proceedings of the Royal Society B, 278: 1158–1164.
Maisano JA, Kearney M, and Rowe T. 2006. Cranial anatomy of the spade-headed amphisbaenian Diplometopon zarudnyi (Squamata, Amphisbaenia) based on high-resolution X-ray computed tomography. Journal of Morphology, 267: 70–102.
Maleev EA. 1956. [Armored dinosaurs of the Upper Cretaceous of Mongolia, Family Ankylosauridae]. Trudy Paleontologicheskogo Instituta, Akademiia Nauk SSSR, 62: 51–91 [In Russian; translation by R. Welch].
Mallon JC, and Anderson JS. 2014a. Implications of beak morphology for the evolutionary paleoecology of the megaherbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology, 394: 29–41.
Mallon JC, and Anderson JS. 2014b. The functional and palaeoecological implications of tooth morphology and wear for the megaherbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. PLoS ONE, 9: e98605.
Mallon JC, Evans DC, Ryan MJ, and Anderson JS. 2013. Feeding height stratification among the herbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. BMC Ecology, 13: 14.
Miyashita T, Arbour VM, Witmer LM, and Currie PJ. 2011. The internal cranial morphology of an armoured dinosaur Euoplocephalus corroborated by X-ray computed tomographic reconstruction. Journal of Anatomy, 219: 661–675.
Nopcsa F. 1915. Die dinosaurier der Siebenbürgischen Landesteile Ungarns [The dinosaurs of the Transylvanian province in Hungary]. Mitteilungen Jahrbuch Ungarische Geologische Reichsanstalt 23: 1–26 [In German; translation by DB Weishampel].
Ősi A, Prondvai E, Mallon J, and Bodor ER. 2017. Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornithischia). Historical Biology, 29: 539–570.
Parks WA. 1924. Dyoplosaurus acutosquameus, a new genus and species of armoured dinosaur, and notes on a skeleton of Prosaurolophus maximus. University of Toronto Studies Geological Series 18. pp. 1–35.
Paul GS. 1997. Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs. In Dinofest international: proceedings of a symposium sponsored by Arizona State University. Edited by DL Wolberg, E Stump, and GD Rosenberg. Academy of Natural Sciences, Philadelphia, Pennsylvania. pp. 129–142.
Penkalski P. 2001. Variation in specimens referred to Euoplocephalus tutus. In The armored dinosaurs. Edited by K Carpenter. Indiana University Press, Bloomington, Indiana. pp. 363–385.
R Core Team. 2015. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria [online]: Available from R-project.org/.
Rieppel O, Kley NJ, and Maisano JA. 2009. Morphology of the skull of the white-nosed blindsnake, Liotyphlops albirostris (Scolecophidia: Anomalepididae). Journal of Morphology, 270: 536–557.
Russell DA, and Manabe M. 2002. Synopsis of the Hell Creek (uppermost Cretaceous) dinosaur assemblage. Geological Society of America Special Paper, 361: 169–176.
Sternberg CM. 1951. Complete skeleton of Leptoceratops gracilis Brown from the Upper Edmonton Member on the Red Deer River, Alberta. Annual Report of the National Museum of Canada Bulletin, 123: 225–255.
Sullivan RM. 1999. Nodocephalosaurus kirtlandensis, gen. et sp. nov., a new ankylosaurid dinosaur (Ornithischia: Ankylosauria) from the Upper Cretaceous Kirtland Formation (Upper Campanian), San Juan Basin, New Mexico. Journal of Vertebrate Paleontology, 19: 126–139.
Vickaryous MK, and Russell AP. 2003. A redescription of the skull of Euoplocephalus tutus (Archosauria: Ornithischia): a foundation for comparative and systematic studies of ankylosaurian dinosaurs. Zoological Journal of the Linnean Society, 137: 157–186.
Vickaryous MK, Russell AP, Currie PJ, and Zhao X-J. 2001. A new ankylosaurid (Dinosauria: Ankylosauria) from the Lower Cretaceous of China, with comments on ankylosaurian relationships. Canadian Journal of Earth Sciences, 38: 1767–1780.
Vickaryous MK, Maryańska T, and Weishampel DB. 2004. Ankylosauria. In The Dinosauria. Edited by DB Weishampel, P Dodson, and H Osmolska. 2nd edition. University of California Press, Berkeley, California. pp. 363–392.
Wilson LE. 2008. Comparative taphonomy and paleoecological reconstruction of two microvertebrate accumulations from the Late Cretaceous Hell Creek Formation (Maastrichtian), eastern Montana. Palaios, 23: 289–297.
Witmer LM, and Ridgely RC. 2008. The paranasal air sinuses of predatory and armored dinosaurs (Archosauria: Theropoda and Ankylosauria) and their contribution to cephalic structure. The Anatomical Record, 291: 1362–1388.
Witmer LM, Sampson SD, and Solounias N. 1999. The proboscis of tapirs (Mammalia: Perissodactyla): a case study in novel narial anatomy. Journal of Zoology, 249: 249–267.
Young MT, Brusatte SL, Ruta M, and de Andrade MB. 2010. The evolution of Metriorhynchoidea (Mesoeucrocodylia, Thalattosuchia): an integrated approach using geometric morphometrics, analysis of disparity, and biomechanics. Zoological Journal of the Linnean Society, 158: 801–859.
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Victoria M. Arbour and Jordan C. Mallon. 2017. Unusual cranial and postcranial anatomy in the archetypal ankylosaur Ankylosaurus magniventris. FACETS.
2(): 764-794. https://doi.org/10.1139/facets-2017-0063
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Dinosaur tracks from the Frontier Formation, Montana: preservation, distribution and palaeoecological significance for the middle Cretaceous terrestrial ecosystems of North America
2. First definitive ankylosaurian dinosaur from the Cretaceous of Jilin Province, northeastern China
3. Living alone or moving in herds? A holistic approach highlights complexity in the social lifestyle of Cretaceous ankylosaurs
4. Osteology and Taxonomy of British Wealden Supergroup (Berriasian–aptian) Ankylosaurs (Ornithischia, Ankylosauria)
5. The braincase of Bissektipelta archibaldi — new insights into endocranial osteology, vasculature, and paleoneurobiology of ankylosaurian dinosaurs
6. Tail Weaponry in Ankylosaurs and Glyptodonts: An Example of a Rare but Strongly Convergent Phenotype
7. Competition structured a Late Cretaceous megaherbivorous dinosaur assemblage
8. The dirty dozen: taxonomical and taphonomical overview of a unique ankylosaurian (Dinosauria: Ornithischia) assemblage from the Santonian Iharkút locality, Hungary
9. The most basal ankylosaurine dinosaur from the Albian–Cenomanian of China, with implications for the evolution of the tail club
10. A “bloat-and-float” taphonomic model best explains the upside-down preservation of ankylosaurs
11. A new southern Laramidian ankylosaurid,
Akainacephalus johnsoni
gen. et sp. nov., from the upper Campanian Kaiparowits Formation of southern Utah, USA