Introduction
Brightness is a key component of primate colour vision; our brain binds together both chromatic and achromatic (brightness) information when interpreting colourful images (
Burns and Shepp 1988;
Croner and Albright 1999;
Clery et al. 2013). Brightness is defined as the attribute of a visual sensation where a stimulus is perceived to be more or less intense, based on the achromatic modulation of photoreceptors by the stimulus (
Wyszecki and Stiles 1982). In the early stages of primate visual processing, chromatic and achromatic information appear to be separated into magnocellular (M) and pavocellular pathways (P) respectively (
Livingstone and Hubel 1988;
Nassi and Callaway 2009). These signals are eventually bound together at a later stage, although where this integration takes place remains unclear and may involve multiple stages (
Nassi and Callaway 2009). Therefore, colour perception incorporating brightness as perceived by primate brains appears to be a complex multi-stage process, and may not be experienced by all animals.
The role of brightness as a visual cue has been previously investigated using behavioural experiments (
Lunau and Maier 1995;
Giurfa et al. 1996;
Kelber 2005) and, therefore, might be an important visual signal in bee-pollinated flowers. Brightness perception is the ability to perceive stimulus intensity differences (
Wyszecki and Stiles 1982;
Reser et al. 2012), where intensity is related to the total amount of energy reflected by a stimulus. To understand whether intensity is a meaningful signal for bee pollinators it is important to test visual perception while isolating other confounding chromatic cues (
Reser et al. 2012). The intensity of flower signals can vary depending on colourless copigments such as flavones, flavanols, or organic acids (
Miller et al. 2011), although flower intensity can also be modulated by physical properties. Such properties include flower thickness, the dense packing of cells via veins or thin cell layers, the curvature in the epidermal layer, and irregularly shaped granules with high refractive indices (
Stavenga and van der Kooi 2016;
van der Kooi et al. 2016,
2017). Ecological studies theoretically investigating brightness often involve pollinator-mediated selection experiments, where selection pressures towards specific colour traits are quantified (
Caruso et al. 2010;
Renoult et al. 2013;
Wassink and Caruso 2013;
Sletvold et al. 2016). Such studies provide some evidence that brightness may be important for pollinators. For example,
Renoult et al. (2013) found that bumble bees drive selection on the brightness component of
Centaurea cyanus Linnaeus flower colouration. However, these experiments do not reveal how bees are processing these components of complex colour signals. This is partially because achromatic visual channels are difficult to isolate from other visual or colour processing mechanisms that can be used by bees (
Giurfa et al. 1996;
Morawetz et al. 2013), and natural spectra may potentially modulate several mechanisms in a highly correlated fashion (
Koethe et al. 2016). In addition,
Renoult et al. (2013) has highlighted a paucity in detection threshold data for achromatic vision in bees. To acquire such data it is important to determine potential detection thresholds to improve our understanding of how bees may first detect the presence of a stimulus against a background, and then, if detection is enabled, how brightness might facilitate the discrimination of a particular target stimulus from an alternative distractor stimulus.
It is surprising that brightness cues in flowers are currently considered important for bees (
Caruso et al. 2010;
Renoult et al. 2013;
Wassink and Caruso 2013;
Sletvold et al. 2016), given the classic psychophysical stance that bees do not process brightness as a dimension of perception in their colour visual system. Early behavioural experiments suggested that honey bees ignore brightness cues in colour choice experiments (
Daumer 1956;
Menzel 1967;
von Helversen 1972). Furthermore, this segregation in visual channels is reflected in the separated use of chromatic or achromatic cues in honey bees at different visual angles (
Giurfa and Vorobyev 1998). Chromatic cues are used when a colour stimulus subtends a large visual angle of approximately >15° (
Giurfa et al. 1996). Alternatively, achromatic cues appear to be used when a stimulus subtends a small angle of approximately 5°–15° (
Giurfa et al. 1996;
Giurfa and Vorobyev 1998), although true achromatic stimuli may be difficult to process (
Giurfa et al. 1996). Bees appear to use achromatic cues at a large visual angle when green contrast is sufficiently high (
Hempel de Ibarra et al. 2000). Therefore, although chromatic and achromatic processing depends on the visual angle subtended by a stimulus to the compound eye, bees may potentially process both these types of information as a combined signal, regardless of the viewing angle.
An additional point to consider is the role of attention and motivation during behavioural experiments. Bees in previous psychophysical studies were not specifically trained to use brightness cues in experiments investigating bee colour vision (
von Frisch 1914;
Menzel 1967). It is, therefore, possible that bees may have ignored brightness in favor of more salient colour cues in these experiments. This is plausible as there seems to be a hierarchy in the bee visual system where some types of chromatic information are weighted above achromatic information (
Morawetz et al. 2013); therefore, it remains unclear whether bees can use brightness in their colour perception when given the proper training. To answer this question, bees need to be trained using a highly motivating appetitive-aversive differential conditioning framework (
Dyer and Neumeyer 2005;
Avarguès-Weber et al. 2010;
Morawetz et al. 2013). This involves the use of both a reward (CS+) and punishment (CS−) while the bee is learning the task (
Dyer and Chittka 2004;
Avarguès-Weber et al. 2010).
In this study, we aimed to expand the current understanding of honey bee brightness processing by investigating whether bees are able to use brightness cues as a component of colour perception when trained using an appetitive-aversive differential conditioning framework; and if so, we sought to establish detection threshold values for brightness perception. We used the well-accepted definitions of brightness (green contrast, and also brightness contrast), as defined by
Spaethe et al. (2001) to reveal whether modulation of brightness over a broad range of values will allow bees to detect achromatic stimuli at a large visual angle.
Discussion
Although brightness and green contrast have been thought to be important cues when considering flower evolution (
Smith et al. 2008;
Hopkins and Rausher 2012;
Renoult et al. 2013;
Sletvold et al. 2016), our findings suggest that honey bees do not make use of such information when using colour vision. Bees were unable to detect the achromatic stimuli from the central region of colour space (
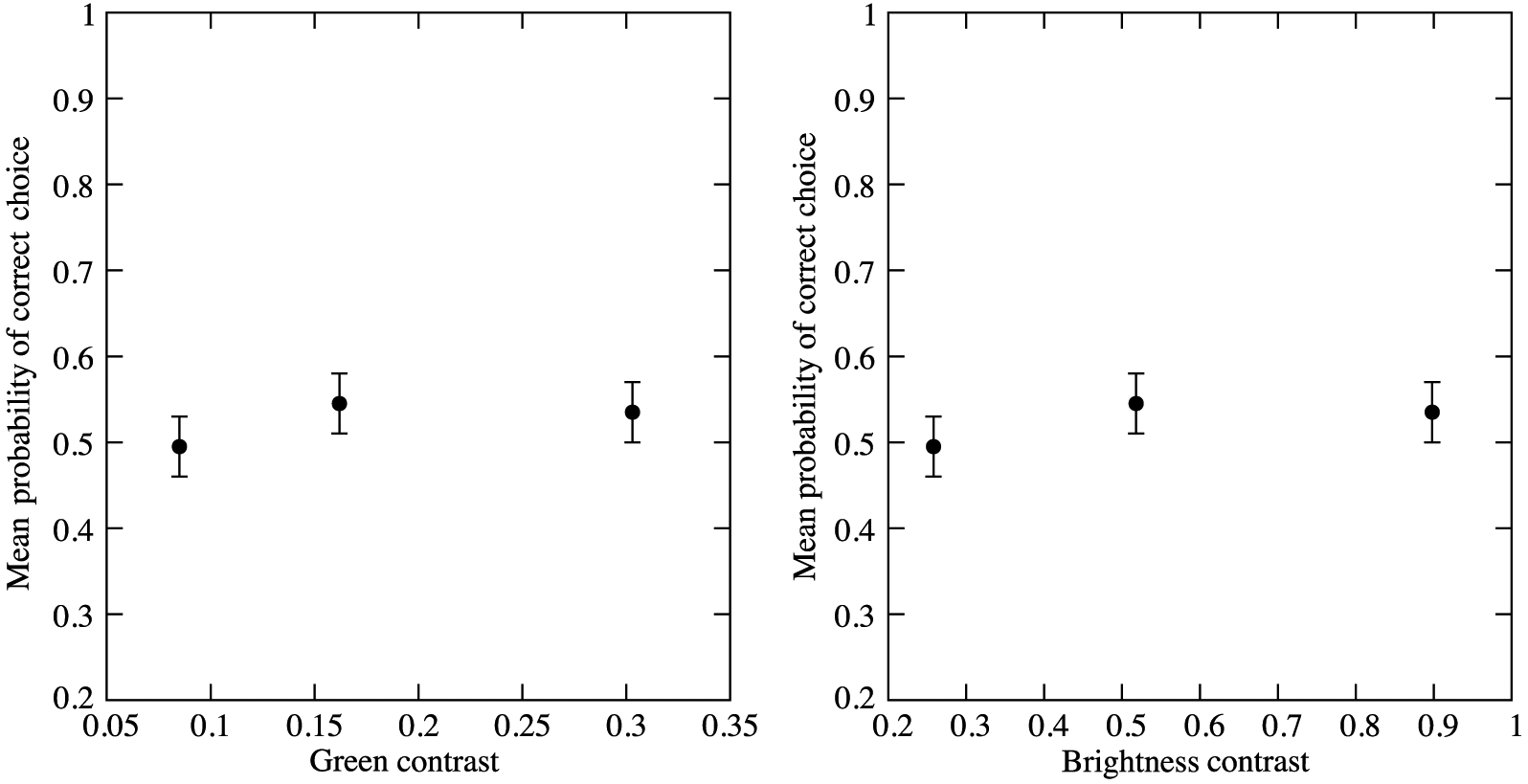
Fig. S4) when presented at a large visual angle that mediates colour processing, even when brightness and green contrast were modulated over a broad range (
Fig. 2). This finding was confirmed following extensive training with the most salient achromatic stimulus (see
Supplementary Material 1). Consequently, we were unable to establish precise detection thresholds of achromatic cues in bee colour vision; it appears that bees do not process brightness information for single target visual processing (
Daumer 1956;
Menzel 1967;
Reser et al. 2012;
van der Kooi et al. 2018). Our findings are consistent with the colour modeling predictions for both the Hexagon and COC models as the stimuli are below the perceptual threshold for honey bees (
Table 2), but our results are not consistent with the RNL (colour) model, which predicted that bees should have been able to process the most salient achromatic stimulus, and the achromatic RNL model actually predicts that all three stimuli should have been easily processed using “brightness” cues. However, although the RNL-achromatic model is based on combined physiological parameters from several vertebrate species, it has not been experimentally validated for bees. We, thus, show in the current study that RNL modeling is not well validated for bee vision considering achromatic processing, and recent work suggests the model is also poor for colour modeling of bee behaviour (
Avarguès-Weber et al. 2010;
Garcia et al. 2017b). Thus, if RNL modeling is to be used for other animals it will be essential to conduct behavioural experiments to validate RNL model predictions. The fact that bees seem to be unable to use achromatic cues in colour tasks agrees with recent findings that flowers do not produce more white signals in low intensity illumination environments (
Binkenstein and Schaefer 2015), and that flowers rarely reflect more than 50% of the incident light, regardless of dominant wavelength, suggesting that higher reflectance does not increase the conspicuousness of the flowers (
van der Kooi et al. 2016). However, bees are tuned to the chromatic signals provided by flowers (
Dyer et al. 2012).
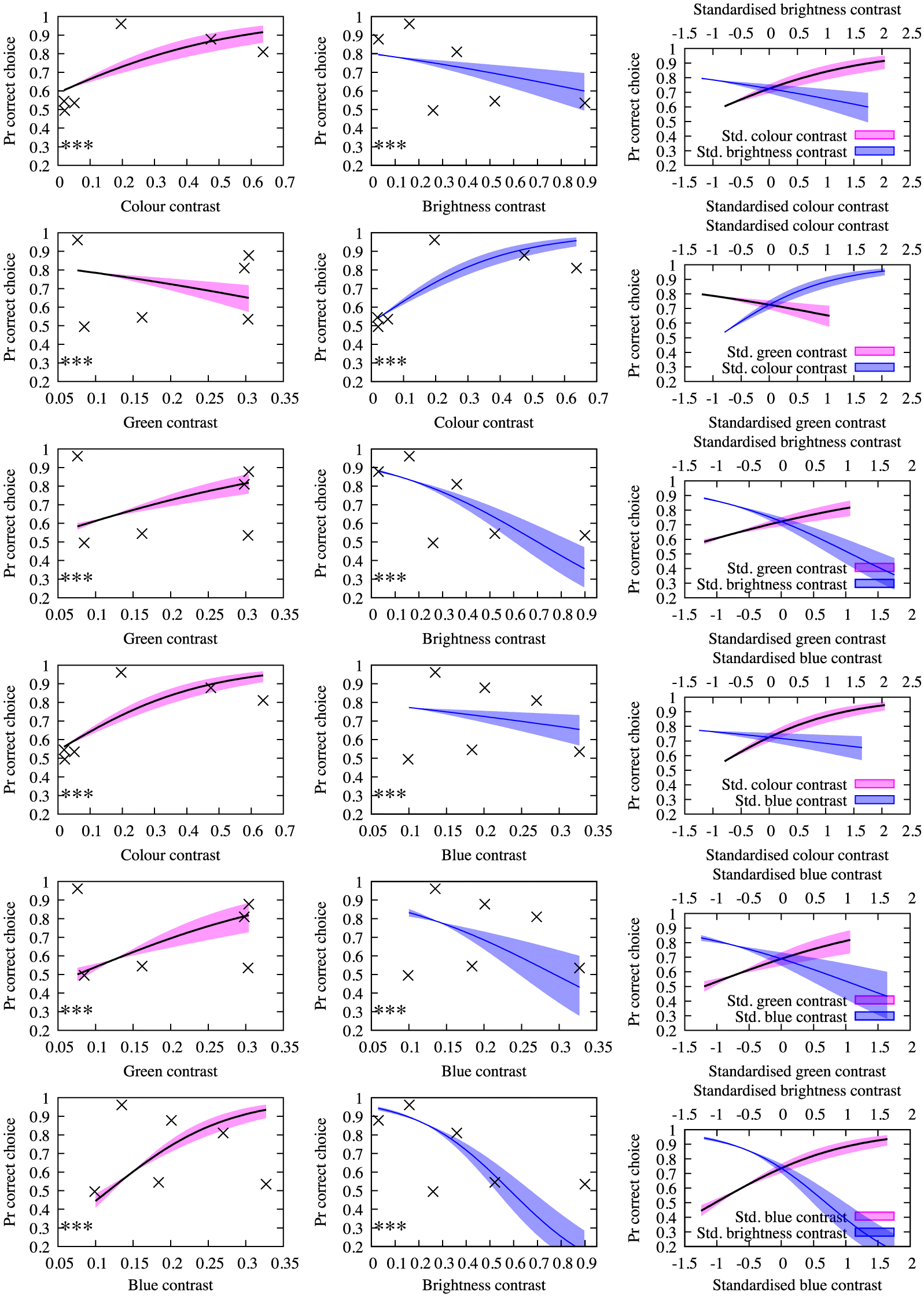
Our findings show that caution is required when investigating pollinator-mediated selection of flower colours, as it is possible that the importance of traits such as brightness may be confounded by their strong relationships with colour when processed by the visual system of an insect pollinator. This is likely the reason why brightness contrast and green contrast were found to be significant main effects in GLMMs where chromatic stimuli were included in the data set (
Models: 2 and
3), as the addition of these chromatic stimuli may be driving the significance of brightness contrast and green contrast as a consequence of being directly correlated with these variables (
Fig. S2). Therefore, it is possible that the individual relationships between variables in
Models 2 and
3 may be biologically meaningless (
Fig. 3). Without considering the psychophysical evidence showing that bees cannot actually detect stimuli when chromatic contrast is low (
Fig. 2), it can be tempting to conclude that brightness and green contrast are both important, as suggested by a simple interpretation of the statistical analysis. Therefore, it is difficult to disentangle the effect of chromatic information from brightness or green contrast when attempting to interpret the importance of chromatic or achromatic traits individually. A qualitative analysis of the main effects from
Models: 3 revealed that potential interactions may exist between all variables of interest. This provides further evidence regarding the complexity of colour as a signal, and the difficulty in disentangling the effects of each factor from the others. These findings call into question whether it is appropriate to investigate these factors individually, as is commonly done in studies exploring flower colours (
Smith et al. 2008;
Caruso et al. 2010;
Hopkins and Rausher 2012;
Renoult et al. 2013;
Wassink and Caruso 2013;
Sletvold et al. 2016). Specifically, colour, by definition, is a construct of an animals’ brain (
Lennie 2000;
Dyer 2012), and quality behavioural data should be carefully considered when attempting to assess colour information processing in animals.
Our findings are supported by early psychophysical and behavioural experiments that suggested that bees ignore brightness cues when processing colour stimuli (
Daumer 1956;
Menzel 1967). They are also consistent with studies investigating the role of visual angle on honey bee colour processing, as bees were unable to detect the achromatic cues at a large visual angle (
Giurfa et al. 1996;
Giurfa and Vorobyev 1998). This inability to process the achromatic cues is likely to be independent of potential attentional confounds, as we trained bees using an appetitive-aversive differential conditioning framework. Therefore, it appears that bees do not process brightness as a dimension of colour perception for single target detection. Achromatic information might only be processed separately for special tasks like motion perception (
von Hess 1913;
Kaiser and Liske 1974;
Stojcev et al. 2011). This conclusion is further supported by neuroanatomical and electrophysiological studies that suggest that brightness and colour information are processed in parallel and independent neural pathways in the bee brain (
Paulk et al. 2008;
Dyer et al. 2011). In the honey bee neuroanatomy, visual information is passed from photoreceptors in the eye to the lamina, the lobulla, and then to higher colour processing areas (
Dyer et al. 2011). The lamina primarily receives input from long-wavelength-sensitive photoreceptors (
Menzel 1974;
Ribi 1975;
Dyer et al. 2011). Alternatively, information received from the short-wavelength-sensitive and medium-wavelength-sensitive photoreceptors are instead passed through the lamina directly to the medulla, and then passed onwards to higher colour processing areas (
Dyer et al. 2011). Despite these independent neural pathways, honey bees are able to detect achromatic signals at large visual angles if green contrast is sufficiently high (e.g., black against white;
Giurfa et al. 1996). As the brightest stimulus from our study had a green contrast of 0.3, bees may be able detect an achromatic signal with a green contrast greater than this, but such stimuli are also likely to modulate chromatic contrast.
Although honey bees may not use brightness cues when viewing flower colours, the green contrast of a flower in the presence of chromatic contrast appears to allow bees to more easily detect it against the background when viewed from a small visual angle (
Giurfa et al. 1996;
Giurfa and Vorobyev 1998;
Bukovac et al. 2017). However,
Giurfa et al. (1996) found that for honey bees, one stimulus (HKS-21N) that lacked chromatic contrast but contained green contrast was poorly processed by bees at a small visual angle, and our control experiment yielded results consistent with this finding (see
Supplementary Material 1). Colour vision is, therefore, mainly used when bees approach a flower and it subtends a large visual angle (
Giurfa et al. 1996). It is also possible that insect pollinators other than bees include brightness as a dimension of colour vision, and it may be these species that drive the selection of brightness cues in flowers. For example, diurnal hawkmoths and
Papilio butterflies have been found to use achromatic cues when landing on or probing artificial flowers (
Koshitaka et al. 2011;
Goyret and Kelber 2012). Further research is, thus, required to elucidate how other insect pollinators process brightness cues in floral displays.
The fact that chromatic and achromatic processing appears to occur independently in honey bees poses an interesting question: Why is this separation more pronounced in bees than in other animals such as primates? For primates, it is plausible that brightness may allow for an improved ability to discriminate between stimuli, especially as interactions between colour and brightness can change the appearance of an object under different viewing conditions (
Xing et al. 2015). Sexual selection and foraging demands that are uniquely experienced by primates are also likely to have influenced the evolution of a colour visual system including brightness as a dimension (
Surridge et al. 2003;
Fernandez and Morris 2007). In contrast, honey bees may have little use for brightness information in a foraging context and, therefore, their colour vision may be sufficient for discriminating between flower species. One possible explanation for this segregation is that the inclusion of brightness information in colour processing may confound its initial purpose of solving the problem of lightness constancy (
von Campenhausen 1986;
Maximov 2000). Achromatic vision has been shown to be less reliable than colour vision under changing light conditions (
Maximov 2000;
Kelber et al. 2003). This is because changes in illumination can result in large variations in receptor signals (
Kelber et al. 2003). Therefore, it has been proposed that the evolution of colour vision was a solution to lightness constancy (
von Campenhausen 1986;
Maximov 2000). Opponency between two spectrally distinct photoreceptors allows for the ratio of light in a scene to be calculated; therefore, the changing signal of background illumination can be differentiated from other changes in the visual scene (
Maximov 2000). If colour vision is indeed a solution to lightness constancy then it would make little sense to feed brightness information into colour processing, as this may result in unreliable output signals. Furthermore, a recent study investigating the dorsal ocelli in honey bees revealed that the ocellar photoreceptors are able to provide information regarding the spectral quality of ambient light conditions to the visual system (
Garcia et al. 2017a). This information is then integrated with colour signals from the frontal compound eyes through a direct neural pathway to allow for a highly accurate reconstruction of flower colour. Therefore, it may not be important to process brightness information captured by the frontal compound eyes, as the required spectral information necessary for solving lightness constancy issues is already provided by the dorsal ocelli.