British Columbia freshwater salmon hatcheries demonstrate minimal contribution to piscine orthoreovirus (PRV) regional occurrence with no evidence for nonendemic strain introductions
Abstract
Piscine orthoreovirus genotype 1 (PRV-1) is a common virus in farmed and wild salmon in the northeastern Pacific Ocean. Its regional occurrence in freshwater is far less clear. From 2019 to 2021, tissues of 5619 juvenile anadromous salmon (primarily Atlantic, Chinook, and coho) sampled from 12 commercial and 27 enhancement British Columbia hatcheries during 83 sampling events were screened for PRV-1 prior to seawater entry. More than 2200 (∼40%) were also screened using a Pan-PRV assay targeting all known PRV genotypes. PRV-1 was detected in four coho salmon at two freshwater enhancement facilities and in one Chinook salmon at a commercial facility. Partial (S1 segment) genome sequencing identified detections to be of the PRV-1 subgenotype endemic to the northeastern Pacific. PRV-1 was not detected (5611 individuals; 99.9%) or test results were inconclusive (3 individuals; 0.05%) for all remaining samples screened for PRV-1. PRV-2 and PRV-3 were not detected using the Pan-PRV assay. It is concluded that commercial and enhancement freshwater hatcheries of British Columbia contribute minimally to the prevalence and persistence of PRV-1 in anadromous salmon of the northeastern Pacific, and these hatcheries appear not to have contracted or participated in the distribution of nonendemic forms of PRV in recent years.
Introduction
Piscine orthoreovirus (PRV) is a common virus of salmon that is endemic (i.e, has maintained constant presence and/or usual prevalence without a need for external introduction) to North America, Europe, South America, and Japan (Polinski et al. 2020). Infections typically manifest as high-load blood infections where red blood cells are the primary target for PRV replication (Wessel et al. 2015). In many instances, fish do not clear infections once infected, and the virus can persist in infected populations at relatively high loads for over a year (Garver et al. 2016). The virus has also been refractory to in vitro culture (Pham et al. 2020), thus requiring molecular methods for diagnosis and load estimations (Palacios et al. 2010; Polinski et al. 2019).
Phylogenetic analysis of PRV has identified three major genotypes: PRV-1, PRV-2, and PRV-3. There are also at least two subgenotypes for PRV-1 (PRV-1a and 1b) and PRV-3 (PRV-3a and 3b) (Siah et al. 2020; Sørensen et al. 2020; Godoy et al. 2021). Genotype occurrence roughly corresponds to discrete hosts and/or regions (Polinski et al. 2020). PRV-1 is found predominately in Atlantic, Chinook, and coho salmon of Europe and the Americas, PRV-2 has only been detected in coho salmon of Japan, and PRV-3 is primarily found in rainbow trout and brown trout in Europe and rainbow trout and coho salmon in Chile (Kibenge et al. 2013; Takano et al. 2016; Cartagena et al. 2020). Separate diagnostic assays that have been developed to detect PRV-1, PRV2, and PRV-3 are not effective for cross-genotype detections. Thus, identification of multiple PRV genotypes within a single fish requires multiple assays to be run, or a recently developed Pan-PRV assay must be utilized (Zhao et al. 2021).
Phylogenetic characterization of PRV-1 has identified regional differentiation within the PRV-1a and PRV-1b subgenotypes, where both subgenotypes co-occur in Norway and Chile, whereas only PRV-1a has been identified in the northwestern Atlantic and northeastern Pacific (Siah et al. 2020). Indeed, PRV-1a endemic to the northeastern Pacific can be differentiated from PRV-1a found in the Atlantic through phylogenetic comparison, either by whole-genome sequencing or individually comparing the S1 genomic segment of the virus (Kibenge et al. 2019; Siah et al. 2020; Godoy et al. 2021). Thus, putative introductions of nonendemic forms of PRV to the northeastern Pacific (either genotype or subgenotype) should be readily identifiable through whole-genome or S1 comparisons.
PRV has sometimes been associated with disease for which variations in pathogenicity can partially be explained by genotype/subgenotype variations (Polinski et al. 2020; Wessel et al. 2020). In Norway, challenge trials using PRV-1b have demonstrated a causal link between PRV-1b and a disease known as heart and skeletal muscle inflammation (HSMI) in farmed Atlantic salmon (Wessel et al. 2017). Interestingly, challenge trials using PRV-1a from the Atlantic have demonstrated less pathogenicity compared with PRV-1b (Wessel et al. 2020), and numerous challenge trials involving PRV-1a in the northeastern Pacific have demonstrated low virulence to both Atlantic and Pacific salmon (Garver et al. 2015, 2016; Polinski et al. 2019, 2021b; Zhang et al. 2019; Purcell et al. 2020). In Japan, PRV-2 is considered a driver of an anemic condition of farmed coho salmon known as erythrocytic inclusion body syndrome (Takano et al. 2016), and PRV-3 causes an HSMI-like condition of rainbow trout in Europe (Vendramin et al. 2019) and jaundice/anemia syndrome in farmed coho salmon in Chile (Cartagena et al. 2020). There may also be an association between PRV-1a and heart inflammation and/or anemia in some instances (Cicco et al. 2017, 2018). Introductions of PRV genotypes/subgenotypes into nonendemic regions could therefore have potential health implications for resident salmon populations.
In the northeastern Pacific, the occurrence of PRV-1a is well documented in ocean stocks of salmon (Polinski et al. 2020). However, occurrence of PRV-1a (or any other genotype/subgenotype) in freshwater juvenile anadromous salmon within the region is far less clear. It is apparent that nearly all net-pen farmed salmon in British Columbia, Canada, become infected with PRV-1 within the first 18 months at sea (Polinski et al. 2022), and cumulative data from multiple wild Pacific salmon surveys have identified an overall regional prevalence in ocean-caught Chinook and coho salmon between 2013 and 2019 of approximately 6% (Polinski et al. 2020). Historically, there has been at least some evidence that PRV-1 has occurred in commercial Canadian salmon hatcheries rearing juvenile fish (Marty et al. 2015; Garver et al. 2016), although distribution and temporal changes in prevalence within freshwater facilities are unknown. There have been no reports for PRV-2 or PRV-3 in the northeastern Pacific, although we are unaware of any studies that have actively targeted either of these genotypes for diagnostics.
In this study, our aims were to (1) define the current regional prevalence of PRV-1 in freshwater anadromous salmon hatchery facilities of western Canada, (2) identify whether alternative genotypes/subgenotypes (e.g., PRV-1b, PRV-2, or PRV-3) are currently occurring within freshwater populations in the region, and (3) confirm interlaboratory agreement for PRV testing between regional laboratories.
Materials and methods
Sample collection
Freshwater anadromous salmon hatcheries in British Columbia can be categorized as either commercial facilities that support seawater net-pen aquaculture, or those that supplement naturally occurring Pacific salmon through the Department of Fisheries and Oceans Canada (DFO) Salmonid Enhancement Program (SEP). SEP hatcheries can be subcategorized into federally operated hatcheries and community-operated hatcheries, the latter either in association with the Community Economic Development Program (CEDP) or Public Involvement Program (PIP). For this study, samples were collected from facilities of each category/subcategory distributed across the geographic breadth of British Columbia (Fig. 1). Kidney and/or spleen tissues were targeted in all sampling events owing to the affinity for PRV to accumulate in these erythrocyte-rich organs (Garver et al. 2016).
Fig. 1.
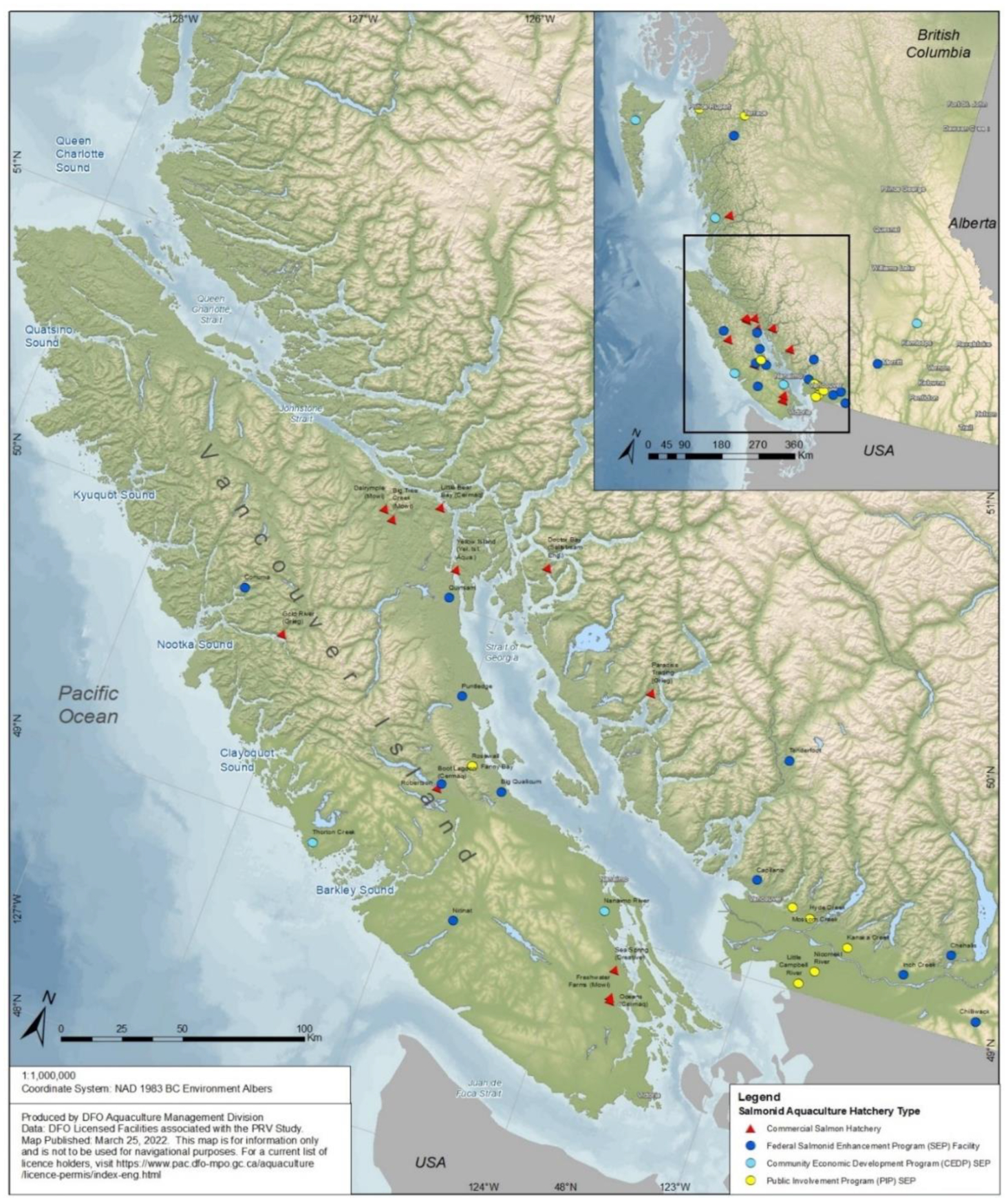
Sampling of commercial hatcheries was conducted as part of the Fish Health Auditing and Surveillance Program performed by DFO Aquaculture Management Division. Sixty fish were targeted in each instance to ensure 95% confidence in identifying at least 5% prevalence of PRV at the population level using the USDA APHIS animal sample size calculator (https://www.aphis.usda.gov) assuming 95% diagnostic test sensitivity based on performance of similar qPCR assays (Polinski et al. 2021a; Yang et al. 2022; Delphino et al. 2023). Fish were sampled from the general population at all locations as neither morbidity nor outward visual appearance/behavior have shown correlation with PRV infection prevalence or load (Takano et al. 2016; Wessel et al. 2017; Polinski et al. 2019, 2022; Marty et al. 2020). Samples were collected at all 12 licensed commercial freshwater salmon facilities in British Columbia: 9 that exclusively rear Atlantic salmon and 3 that exclusively rear Chinook salmon (Supplemental material 1). Each facility was visited and sampled 2–6 times between August 2019 and September 2021 culminating in a total of 53 sampling events (Fig. 2; Supplemental material 1). In each instance, kidney samples (5–50 mg) were collected from 60 fish per sampling event, placed into individual 2 mL microtubes containing 1 mL of RNAlater® nucleic acid preservation solution and stored at −20 °C. Frozen samples were transported to the DFO Pacific Biological Station (DFO-PBS) in Nanaimo, British Columbia, for processing.
Fig. 2.
Of the more than 50 SEP freshwater facilities operating in British Columbia, 27 were opportunistically sampled during 39 sampling events between March 2020 and June 2021 (Fig. 2). This included 14 Federal, 5 CEDP, and 8 PIP facilities based on availability and program interest. Facilities encompassed the full geographic range of SEP in British Columbia (Fig. 1; Supplemental material 1) and facilities containing Chinook and coho salmon were specifically targeted because (i) overall prevalence of PRV-1 in the northeastern Pacific has been documented to be highest in these species relative to other Pacific salmonids (Purcell et al. 2018; Polinski et al. 2020) and (ii) PRV-2 and PRV-3 have also been documented in coho salmon in freshwater from other countries (Takano et al. 2016; Cartagena et al. 2020). Samples were collected as close as possible to the end of a production cycle (typically within 1 month of release) to maximize the likelihood of detecting infections that are known to persist in aquaculture-associated populations for at least 14 months once established (Garver et al. 2016).
All SEP sampling was conducted either by DFO researchers during hatchery visits, or whole fish were shipped directly to DFO-PBS by hatchery personnel. On-site tissue collections by DFO researchers were performed during 15 sampling events at 14 SEP facilities. Tissues (8–22 mg kidney and/or spleen; N = 60 per hatchery sampling event) were collected from MS-222 euthanized fish, individually weighed in 1.2 mL microtubes, and immediately frozen at −80 °C. During five additional sampling events involving five hatcheries, MS-222 euthanized whole fish were collected on-site by DFO researchers and immediately frozen at −80 °C for transport. In the laboratory, specimens were thawed at 4 °C, and kidney and/or spleen tissues excised, weighed, and processed for nucleic acid extraction as described below. For the remaining 19 sampling events conducted across 17 facilities, MS-222 euthanized fish were collected whole by hatchery personnel into bags based on species and stock of origin. Fish were either immediately shipped on ice (1 event) or frozen at −20 °C (18 events) and then shipped to PBS, whereupon fish were thawed at 4 °C and tissues collected as described above.
Nucleic acid extraction and RT-qPCR screening
Nucleic acid extraction and diagnostic detection of PRV RNA was conducted at either the DFO-PBS (5219 samples), the British Columbia Centre for Aquatic Health Sciences (CAHS; 200 samples), or the Okanagan Nation Alliance Fisheries Research and Diagnostic facility (ONA; 200 samples). At DFO-PBS, nucleic acids were purified from tissues either manually using Trizol® (Thermo-Fisher Scientific; 448 samples) or via a semiautomated process involving a Kingfisher Flex and MagMAX™ Pathogen RNA/DNA kit (Thermo-Fisher Scientific; 4771 samples). Eluted RNA (up to 1 µg) from either method was then reverse transcribed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) and screened for PRV-1 (Polinski et al. 2022). A detailed protocol for nucleic acid extraction, reverse transcription, and PRV-1 qPCR screening is provided in Supplementary material 2. A portion of extracted RNA from a subset of samples (2205 samples encompassing all five salmon species and all five subcategories of freshwater facilities; Supplementary material 1) was additionally screened for PRV-1, -2, and -3 using a Pan-PRV assay as previously described (Zhao et al. 2021). Absolute PRV quantification was determined in each instance by serial dilution of a 482 bp double-stranded DNA gBLOCK fragment (Integrated DNA Technologies) consisting of sequence targeted by the qPCR primer and probe (Garver et al. 2016; Zhao et al. 2021). A six-step 10-fold dilution series of the gBLOCK fragment spanning a dynamic range of 100–107 target copies per reaction was incorporated in duplicate into each run.
Sample screening conducted at CAHS was performed using the same methods as presented for DFO-PBS (200 samples via MagMAX™ extraction; Supplementary material 1). At ONA, nucleic acid was purified from tissues using Quick-DNA/RNA Pathogen Mag Bead extraction kit (Zymo Research) according to the manufacturer's recommendations. Eluted RNA was reverse transcribed using Quantitech™ reverse transcription kit (Qiagen) and screened for PRV1 qPCR using QuantiNova Probe PCR kit (Qiagen). A detailed protocol for nucleic acid extraction, reverse transcription, and PRV-1 qPCR screening is provided in Supplementary material 2.
Reproducibility for PRV-1 detection
Consistency in PRV-1 detection between laboratories and processing methods was determined by comparing results of replicate kidney samples from 10 farmed Atlantic salmon with predetermined PRV status (7 positive, 3 negative) tested at both DFO and CAHS laboratories. A replicate kidney sample was sent to both laboratories and all sample processing was completed blinded to prescreening PRV status. Additionally, the 10 samples sent to DFO-PBS were divided prior to nucleic acid extraction and processed separately using both the Trizol® or MagMAX™ nucleic acid extraction protocols. To further confirm interlaboratory agreement, population sets (n = 60 per set) from each of 10 commercial hatchery sampling events were equally distributed and screened at DFO, CAHS, or ONA diagnostic laboratories (i.e., n = 20 samples per laboratory per population set; 600 total samples; Supplementary material 1).
S1 genome sequencing and phylogenetic comparisons
S1 genome sequencing was attempted in the four instances in this study where PRV genomic material was detected at a mean concentration greater than 1000 copies/mg (i.e., Ct ∼ <32), which in our experience represents a minimum concentration necessary for reliable sequence coverage using our methods (data not shown). A portion (1 µg) of purified total RNA not used for qPCR analysis was preamplified using primer pairs designed in Primal Scheme (http://primal.zibraproject.org) to target the S1 sequence of PRV-1 (Supplementary material 1) and run in multiplex qPCR, cleaned and sequenced following the protocol outlined by Quick et al. (2017) using a nanopore MinION sequencer. Reads were assembled against the published PRV-1 genome (GenBank GCA_002829625.1; Kibenge et al. 2013) using NanoPipe (Shabardina et al. 2019). Phylogenetically comparisons of S1 segments were performed against a select subset of 230 published S1 sequences previously evaluated for phylogeny (Siah et al. 2020; Supplementary material 1) via a Tamura–Nei neighbor-joining method following Clustal Omega maximum-likelihood alignment using Geneious Prime 2022.1.1.
Results
PRV diagnostic detection
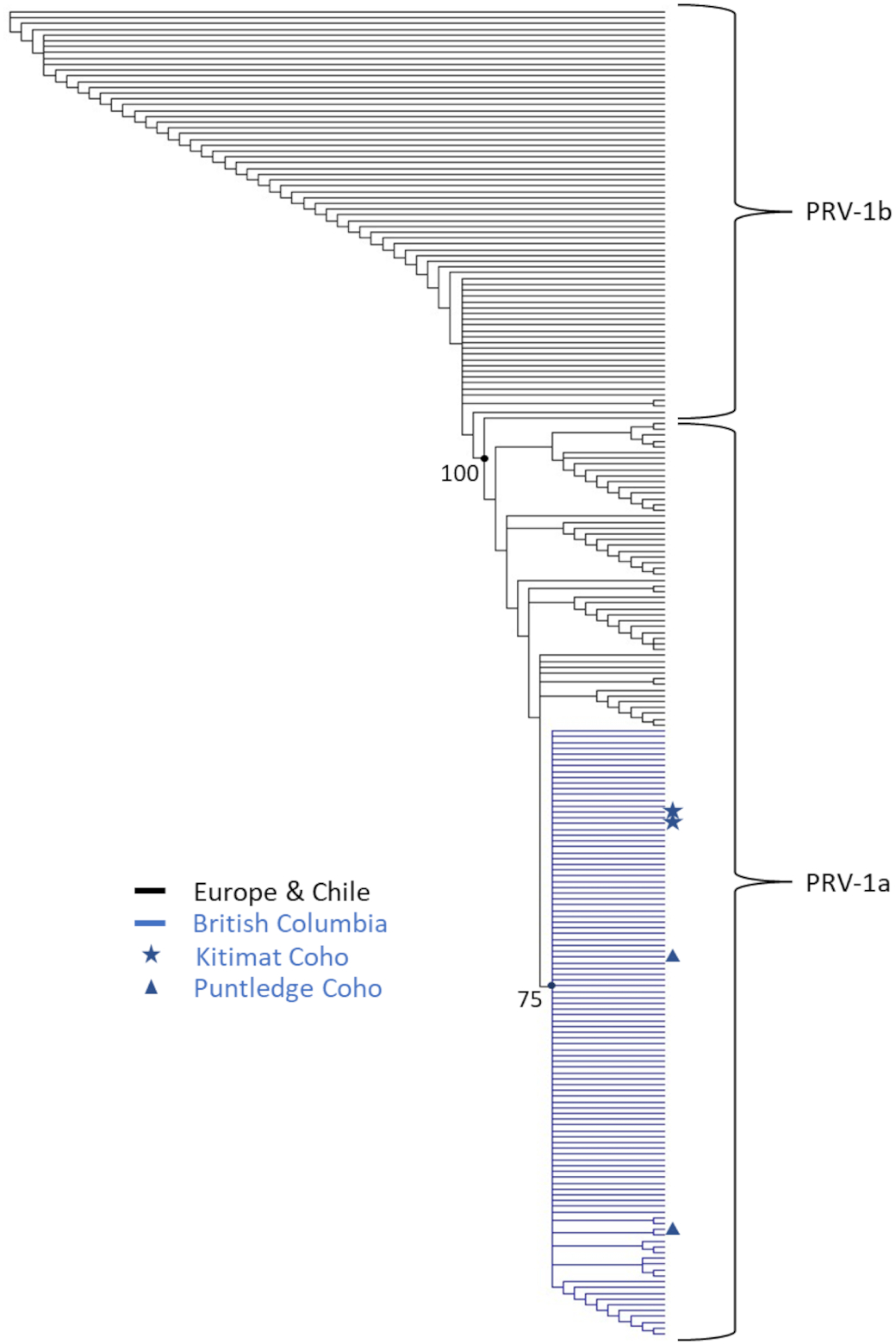
PRV-1 RNA was detected in less than 0.09% of hatchery-reared juvenile salmon sampled in this study. Four coho salmon sampled from two separate federal SEP facilities in 2020 were positive for PRV-1: 2 of 60 fish (3.3%) from Kitimat River hatchery on the north-central coast and 2 of 60 fish (3.3%) from the Puntledge River hatchery located along the northeastern portion of Vancouver Island (Table 1; Supplementary material 1). Subsequent sampling conducted at these facilities in 2021 did not detect PRV-1, and cumulative PRV-1 prevalence for all sampled coho salmon between 2020 and 2021 was 0.2% (4/1920). There were no detections of PRV-1 in commercial facilities raising Atlantic salmon, nor in SEP populations of Chinook, sockeye, or steelhead salmon incorporated into this study (Tables 1 and 2).
Table 1.
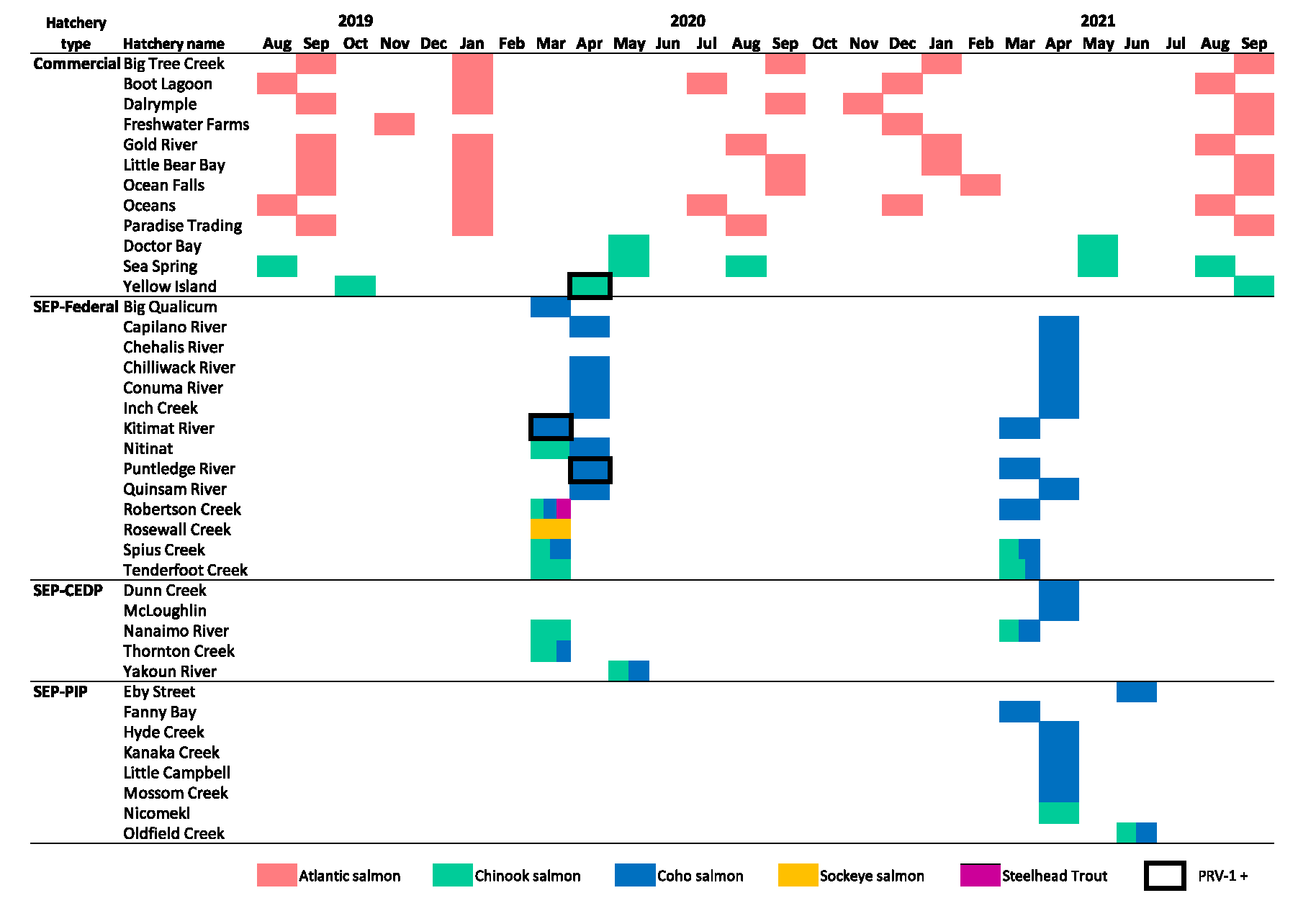
Note: Relative proportions of samples for each species (colored) as well as which sampling events contained PRV-1 detections (boxed) are provided.
Table 2.
| PRV-1 diagnostic result (detection/inconclusive/no detection) | ||||||
|---|---|---|---|---|---|---|
| Private commercial hatcheries | Public enhancement hatcheries | |||||
| Sample type | Atlantic | Chinook | Chinook | Coho | Sockeye | Steelhead |
| Individual fish | 0/1/2516 | 1/1/534 | 0/0/502 | 4/1/1920 | 0/0/60 | 0/0/20 |
| Facilities | 0/1/8 | 1/0/3 | 0/0/9 | 2/0/22 | 0/0/1 | 0/0/1 |
| Events | 0/1/41 | 1/0/8 | 0/0/12 | 2/0/31 | 0/0/1 | 0/0/1 |
In one sampling event in 2020 at Yellow Island commercial Chinook salmon hatchery, 2 of 60 fish were identified with inconclusive PRV-1 screening results. In both instances (one tested at CAHS and one at ONA), low copy number detections (Ct > 35 indicative of putative PRV RNA concentrations below 100 copies/mg) were identified in one of two technical qPCR replicates in each instance. Repeated testing of the CAHS sample yielded further inconclusive results and the extracted RNA was sent to DFO-PBS for confirmatory testing. Testing of the suspect sample at DFO-PBS using the PRV-1 specific assay showed no detection in either technical replicate; however, screening conducted using the Pan-PRV assay (which targets an alternative genomic segment) identified PRV in both technical replicates, albeit at a mean Ct > 35 (<100 copies per mg). We report this as a positive PRV-1 detection even though confirmatory viral sequence analysis was not completed due to insufficient PRV genomic material. The inconclusive result from ONA facility was not retested and PRV-1 was not detected at the Yellow Island facility during 2019 or 2021 samplings, nor was it detected in any other commercial Chinook facility.
There were two additional instances (one at a commercial Atlantic salmon hatchery and one at an SEP federal hatchery with a population of coho), where inconclusive PRV-1 detections occurred with one of two technical qPCR replicates identifying PRV-1 RNA Ct > 35 (<100 copies per mg). Retesting of these samples did not result in positive detections.
Screening 2205 of the collected samples (39.2% of total samples screened for PRV-1) using the Pan-PRV assay—which included samples of each species and from each hatchery category/subcategory—did not detect PRV RNA except in the instances where PRV-1 was detected (or inconclusively detected as described above) using the PRV-1 specific assay. Therefore, there were no indications for PRV-2 or PRV-3 RNA to be present in any of the samples screened using the Pan-PRV assay and PRV-2 or PRV-3 specific assays were not performed.
Phylogenetic comparison of PRV detections
S1 phylogeny of PRV-1 RNA recovered from the four qPCR-positive coho salmon in comparison with 226 previously published sequences identified that all four fish were infected with the subgenotype of PRV-1 (PRV-1a as described by Kibenge et al. (2013) or Group L as described by Siah et al. (2020)) that is endemic to the northeastern Pacific (Fig. 2). The two S1 sequences obtained from Kitimat hatchery in 2020 were identical. The two S1 sequences obtained from Puntledge hatchery in 2020 were slightly different from those identified in Kitimat (2–5 nucleotide substitutions within the 933 bp fragment), but interestingly were also different from each other (7 nucleotide substitutions).
Reproducibility for PRV-1 detection
Detection agreement in classifying PRV-1 status for 10 fish screened at both DFO and CAHS laboratories in this study was 100%. Each lab congruently found seven of the fish to be positive and three to be negative for PRV-1. Relative quantity estimations for positive fish within that population dataset were highly similar between laboratories (Fig. 3a). Interlaboratory agreement was also identical for assessing PRV status at a population level in 9 out of the 10 hatchery sample sets screened across the three labs. The only instance of nonagreement was for the Yellow Island Chinook hatchery samples collected in 2020 (n = 60), where CAHS had 1 inconclusive detection in 20 samples (which DFO-PBS later confirmed to be positive using the Pan-PRV assay as described above), ONA had 1 inconclusive detection in 20 samples, and DFO-PBS had no detections in 20 samples.
Fig. 3.
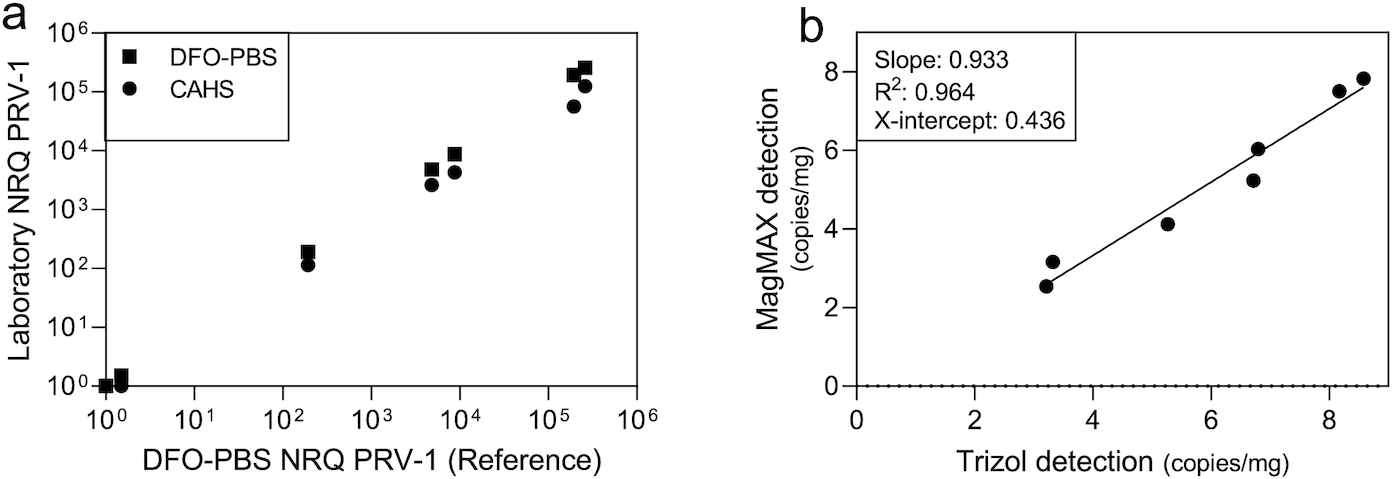
Comparison of the two nucleic acid extraction methods (Trizol vs MagMAX) employed by DFO-PBS and CAHS during the screening process identified high quantitative agreement for PRV-1 detection between the two methods (Fig. 3b). The theoretical limit of detection using the MagMAX protocol was approximately 60 copies/mg from a 10 mg sample, which was slightly lower than when using the Trizol protocol (220 copies/mg from a 10 mg sample) owing to less overall dilution of starting material. However, efficiency for recovering PRV RNA using MagMAX was less than when using Trizol (mean 22 ± 21% SD), thereby making the effective limit of detection for both assays highly similar—approximately 200–300 copies/mg for a 10 mg sample.
Discussion
Transmission of PRV (all genotypes) has been readily accomplished in freshwater and has been well documented in freshwater facilities raising juvenile anadromous salmon from North America, Europe, and Japan (reviewed by Polinski et al. 2020). This has included multiple commercial freshwater Atlantic salmon facilities in British Columbia within the past decade (Marty et al. 2015; Garver et al. 2016; Polinski et al. 2019). Following initial detection of PRV-1 in 2013, commercial Atlantic salmon growers in British Columbia have made concerted efforts toward removing PRV-1 from freshwater facilities. A decreasing trend for PRV-1 prevalence was observed from 2013 to 2018 where 42% of 48 fish sampled from two hatcheries were PRV-1 positive in 2013 and by 2018, 0.2% of 586 fish sampled from eight hatcheries were positive for PRV-1 (Mimeault et al. 2019). Our findings indicate that this decreasing prevalence trend has continued, and that by 2019 all Atlantic salmon facilities have likely been PRV-1 free. It therefore does not appear that freshwater Atlantic salmon facilities used to seed regional seawater net-pens have played a meaningful role in PRV-1 persistence in the region in recent years even though they may have acted as a reservoir historically within the region. Further, given that some Atlantic salmon producers rely on PRV-1 infected broodstock for gametes (Polinski et al. 2022), it also appears that current iodine-based disinfection strategies employed by the industry are effective at deterring egg-associated transmission of PRV-1 as previously suggested (Polinski et al. 2020).
In considering the three commercial Chinook salmon freshwater facilities in British Columbia, this study identified only one individual in one cohort at one facility to be putatively infected with PRV-1. If this detection represents a true positive, it is interesting that this did not appear to create a persistent viral presence in the facility. Laboratory challenge studies with PRV-1 have identified that the virus quickly spreads through a confined population once established, which results in readily detectable persistent infections (Garver et al. 2016; Wessel et al. 2017). Yet, repeat sampling of this facility with a subsequent cohort in 2021 did not detect virus. It is therefore possible that the 2020 detection represents a false positive result. Although qPCR has demonstrated both high diagnostic specificity and sensitivity—often greater than 98% specificity in stringently controlled laboratories (Polinski et al. 2021a; Yang et al. 2022; Delphino et al. 2023)—false positive detections become increasingly likely as sample size increases. Further, false positive detections are also most likely to manifest at or near the lower limit of detection of the qPCR assay due to a combination of increased probability for low-quantity aerosol contamination to occur or nonspecific fluorescence changes to manifest in late qPCR cycles (Polinski et al. 2021a; Meyers and Hickey 2022). In this study, all three inconclusive detections (1 coho, 1 Chinook, and 1 Atlantic) as well as one positive detection from a Chinook salmon were at or below the lower effective limit of detection of the qPCR assay (i.e., less than 1–3 theoretical copies per reaction; Ct > 35) and represent <0.08% of screened samples, implying that theoretical diagnostic specificity was 99.92% or greater in this study even if these detections were false. It therefore becomes highly speculative in defining inconclusive samples from this study as either infected or not infected with PRV-1, and we therefore make no recommendation for dichotomizing inconclusive results presented here. We further treat the one commercial chinook salmon detection in this instance with substantial uncertainty. Irrespective of the validity of the single chinook detection however, it appears that commercial freshwater Chinook salmon facilities have not been a major contributor to the regional prevalence of PRV-1 within the past few years.
The four coho salmon with PRV-1 infections from SEP facilities in this study all had relatively high viral RNA copy numbers. This, in concert with identifying repeat positive samples at the population level in both premises where PRV-1 was detected and ability to sequence a second viral genomic segment, provides strong evidence for diagnostic accuracy in these instances. Interestingly, the relatively low prevalence for PRV-1 in each of the two sample sets (both 3.3%) suggests that viral transmission within each facility was either very low or the populations had only been recently infected. Cumulative regional prevalence of PRV-1 in marine wild coho salmon has been around 6% (Polinski et al. 2020) and PRV-1 has been occasionally detected in returning adults to both facilities (S. Johnson (personal communication)), which provides at least one putative infection source. It was also anecdotally noted that detections in 2020 at Kitimat hatchery occurred shortly after fish were transitioned from well water to river water prior to release. The fact that the S1 sequences from the two positive detections at Puntledge hatchery in 2020 were not identical indicates that perhaps multiple infection sources were present at that facility. Whatever the potential source, it appeared to be transitory, as there was no carrying over of infection between 2020 and 2021 year-classes at either facility. Additionally, the low PRV-1 prevalence in these two instances and the lack of detection at all other SEP facilities indicate that the SEP program at large does not appear a major reservoir for PRV-1 in the region. Historic contributions of PRV also seem unlikely from these facilities given that PRV has not been cleared from a local region/facility to our knowledge once it has been introduced without targeted mitigation and monitoring, and SEP facilities in British Columbia have neither monitored nor actively mitigated against PRV transmission.
The absence of PRV-2 and PRV-3 detection within SEP coho during this study is also significant. Both genotypes have been associated with freshwater coho populations in all other regions where the genotypes are endemic (reviewed by Polinski et al. 2020) and its lack of detection here provides strong evidence that neither of these PRV genotypes are present in the region. However, there is a tropism for PRV-3 to infect rainbow trout in Europe and rainbow trout have a potential ability to clear PRV-3 within 2 or 3 months postinfection (Hauge et al. 2017; Vendramin et al. 2019)—a unique circumstance for PRV–salmon interactions. A current lack of screening for PRV-3 in rainbow/steelhead trout in the northeastern Pacific (only 20 steelhead from one facility were screened in this study) thus provides a potential unexplored reservoir for PRV-3 occurrence in the region, although the fact that both steelhead and coho are sometimes reared concurrently in SEP hatcheries makes it seem unlikely that a putative unidentified PRV-3 rainbow/steelhead trout reservoir has not spilled over into cohabitating coho.
To efficiently screen the more than 5000 samples collected in this study, we developed and assessed a semiautomated process for purifying PRV nucleic acid (MagMAX kits using the Kingfisher Flex). This method proved highly similar in sensitivity and performance to previously published detection methods using phenol/guanidine-based methods (Trizol). Although not directly compared in this study, it also appears that quantitative estimations by our new method are reasonably similar to column-based purification procedures (e.g., QIAGEN's QIAamp Viral RNA mini kit) (Wessel et al. 2020). We anticipate that this method will prove highly desirable in future efforts for screening large population sets for PRV. This may be of particular importance if any of the three PRV genotypes or subgenotypes become nationally or internationally regulated in future.
In conclusion, data generated in this study indicate that commercial and enhancement freshwater salmon facilities of British Columbia currently contribute minimally to the regional ubiquitous seawater prevalence and persistence of PRV-1 and confirm that seawater reservoirs are the primary mechanism for continued PRV-1 persistence in the region. Freshwater facilities also appear not to have contracted or participated in the distribution of nonendemic forms of PRV as neither PRV-2, PRV-3, or Atlantic variants of PRV-1 were detected. Cumulatively, this would indicate that continuation of current aquaculture management strategies and practices in both commercial and SEP freshwater facilities is unlikely to affect regional prevalence of this virus.
Acknowledgements
The authors would like to thank DFO's aquaculture management division, specifically Howie Manchester and his crew for collecting samples during commercial facility audits, as well as Chris O'Grady for producing Fig. 1. The authors also thank Stewart Johnson and Lenora Turcotte for aiding in PRV-1 sequencing and assembly, Christy Thompson for aiding with sample collection at federal SEP facilities, and all SEP hatchery personnel and regional managers who facilitated and allowed sample collections to be conducted at their facilities.
References
Cartagena J., Jiménez C., Spencer E. 2020. Detection of Piscine orthoreoviruses (PRV-1b AND PRV-3a) in farmed Coho salmon with jaundice syndrome from Chile. Aquaculture, 528: 735480.
Cicco E.D., Ferguson H.W., Schulze A.D., Kaukinen K.H., Li S., Vanderstichel R., et al. 2017. Heart and skeletal muscle inflammation (HSMI) disease diagnosed on a British Columbia salmon farm through a longitudinal farm study. PLoS ONE, 12: e0171471.
Cicco E.D., Ferguson H.W., Kaukinen K.H., Schulze A.D., Li S., Tabata A., et al. 2018. The same strain of Piscine orthoreovirus (PRV-1) is involved in the development of different, but related, diseases in Atlantic and Pacific Salmon in British Columbia. FACETS, 3: 599–641.
Delphino M.K., O'Brien N., Laurin E., Whelan D., Burnley H., Hammell K.L., Thakur K.K. 2023. Bayesian analysis of diagnostic sensitivity and specificity for detecting infectious salmon anaemia virus (ISAV) using IFAT and real-time RT-PCR testing from laboratories in Atlantic Canada. Aquaculture, 563: 739006.
Garver K.A., Marty G.D., Cockburn S.N., Richard J., Hawley L.M., Müller A., et al. 2015. Piscine reovirus, but not jaundice syndrome, was transmissible to Chinook salmon, Oncorhynchus tshawytscha (Walbaum), sockeye salmon, Oncorhynchus nerka (Walbaum), and Atlantic salmon, Salmo salar L. Journal of Fish Diseases, 39: 117–128.
Garver K.A., Johnson S.C., Polinski M.P., Bradshaw J.C., Marty G.D., Snyman H.N., et al. 2016. Piscine orthoreovirus from western North America is transmissible to Atlantic salmon and Sockeye salmon but fails to cause heart and skeletal muscle inflammation. PLoS ONE, 11: e0146229.
Godoy M., Medina D.A., Suarez R., Valenzuela S., Romero J., Kibenge M., et al. 2021. Extensive phylogenetic analysis of Piscine orthoreovirus genomic sequences shows the robustness of subgenotype classification. Pathogens, 10: 41.
Hauge H., Vendramin N., Taksdal T., Olsen A.B., Wessel Ø., Mikkelsen S.S., et al. 2017. Infection experiments with novel Piscine orthoreovirus from rainbow trout (Oncorhynchus mykiss) in salmonids. PLoS ONE, 12: e0180293.
Kibenge M.J.T., Iwamoto T., Wang Y., Morton A., Godoy M.G., Kibenge F.S. 2013. Whole-genome analysis of piscine reovirus (PRV) shows PRV represents a new genus in family Reoviridae and its genome segment S1 sequences group it into two separate sub-genotypes. Virology Journal, 10: 10–230.
Kibenge M.J.T., Wang Y., Gayeski N., Morton A., Beardslee K., Mcmillan B., Kibenge F.S.B. 2019. Piscine orthoreovirus sequences in escaped farmed Atlantic salmon in Washington and British Columbia. Virology Journal, 16: 41.
Marty G.D., Morrison D.B., Bidulka J., Joseph T., Siah A. 2015. Piscine reovirus in wild and farmed salmonids in British Columbia, Canada: 1974–2013. Journal of Fish Diseases, 38: 713–728.
Marty G.D., Bidulka J., Joseph T. 2020. Cross-sectional study of histopathology and piscine orthoreovirus during a marine production cycle of farmed Atlantic salmon (Salmo salar L.) in British Columbia, Canada. Journal of Fish Diseases, 43: 1019–1028.
Meyers T.R., Hickey N. 2022. A perspective: molecular detections of new agents in finfish—interpreting biological significance for fish health management. Journal of Aquatic Animal Health, 34: 47–57.
Mimeault C., Polinski M.P., Garver K.A., Jones S.R.M., Johnson S., Boily F., et al. 2019. Assessment of the risk to Fraser River Sockeye Salmon due to piscine orthoreovirus (PRV) transfer from Atlantic Salmon farms in the Discovery Islands area, British Columbia. Canadian Science Advisory Secretariat (CSAS), Research Document 2019/036.
Palacios G., Lovoll M., Tengs T., Hornig M., Hutchison S., Hui J., et al. 2010. Heart and skeletal muscle inflammation of farmed salmon is associated with infection with a novel reovirus. PLoS ONE, 5: e11487.
Pham P.H., Misk E., Papazotos F., Jones G., Polinski M.P., Contador E., et al. 2020. Screening of fish cell lines for piscine orthoreovirus-1 (PRV-1) amplification: identification of the non-supportive PRV-1 invitrome. Pathogens, 9.
Polinski M.P., Marty G.D., Snyman H.N., Garver K.A. 2019. Piscine othoreovirus demonstrates high infectivity but low virulence in Atlantic salmon of Pacific Canada. Scientific Reports, 9.
Polinski M.P., Vendramin N., Cuenca A., Garver K.A. 2020. Piscine orthoreovirus: biology and distribution in farmed and wild fish. Journal of Fish Diseases, 43: 1331–1352.
Polinski M.P., Laurin E., Delphino M.K., Lowe G.J., Meyer G.R., Abbott C.L. 2021a. Evaluation of histopathology, PCR, and qPCR to detect Mikrocytos mackini in oysters Crassostrea gigas using Bayesian latent class analysis. Diseases of Aquatic Organisms, 144: 21–31.
Polinski M.P., Zhang Y., Morrison P.R., Marty G.D., Brauner C.J., Farrell A.P., Garver K.A. 2021b. Innate antiviral defense demonstrates high energetic efficiency in a bony fish. BMC Biology, 19: 1–15.
Polinski M.P., Gross L., Marty G.D., Garver K.A. 2022. Heart inflammation and piscine orthoreovirus genotype-1 in Pacific Canada Atlantic salmon net-pen farms: 2016-2019. BMC Veterinary Research, 18: 1–14.
Purcell M.K., Powers R.L., Evered J., Kerwin J., Meyers T.R., Stewart B., Winton J.R. 2018. Molecular testing of adult Pacific salmon and trout (Oncorhynchus spp.) for several RNA viruses demonstrates widespread distribution of piscine orthoreovirus in Alaska and Washington. Journal of Fish Diseases, 41: 347–355.
Purcell M.K., Powers R.L., Taksda T., Mckenney D., Conway C.M., Elliott D.G., et al. 2020. Consequences of Piscine orthoreovirus genotype one (PRV-1) infections in Chinook salmon (Oncorhynchus tshawytscha), coho salmon (O. kisutch) and rainbow trout (O. mykiss). Journal of Fish Diseases, 43: 719–728.
Quick J., Grubaugh N.D., Pullan S.T., Claro I.M., Smith A.D., Gangavarapu K., et al. 2017. Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nature Protocols, 12: 1261–1276.
Shabardina V., Kischka T., Manske F., Grundmann N., Frith M.C., Suzuki Y., Makałowski W. 2019. NanoPipe: a web server for nanopore MinION sequencing data analysis. GigaScience, 8: giy169.
Siah A., Breyta R., Warheit K., Gagne N., Purcell M.K., Morrison D., et al. 2020. Genomes reveal genetic diversity of Piscine orthoreovirus in farmed and free-ranging salmonids from Canada and USA. Virus Evolution, 6: veaa054.
Sørensen J., Vendramin N., Priess C., Kannimuthu D., Henriksen N.H., Iburg T.M., et al. 2020. Emergence and spread of piscine orthoreovirus genotype 3. Pathogens, 9: 823.
Takano T., Nawata A., Sakai T., Matsuyama T., Ito T., Kurita J., et al. 2016. Full-genome sequencing and confirmation of the causative agent of erythrocytic inclusion body syndrome in coho salmon identifies a new type of piscine orthoreovirus. PLoS ONE, 11: e0165424.
Vendramin N., Kannimuthu D., Olsen A.B., Cuenca A., Teige L.H., Wessel Ø., et al. 2019. Piscine orthoreovirus subtype 3 (PRV-3) causes heart inflammation in rainbow trout (Oncorhynchus mykiss). Veterinary Research, 50: 14.
Wessel Ø., Olsen C.M., Rimstad E., Dahle M.K. 2015. Piscine orthoreovirus (PRV) replicates in Atlantic salmon (Salmo salar L.) erythrocytes ex vivo. Veterinary Research, 46: 26.
Wessel Ø., Braaen S., Alarcon M., Haatveit H., Roos N., Markussen T., et al. 2017. Infection with purified Piscine orthoreovirus demonstrates a causal relationship with heart and skeletal muscle inflammation in Atlantic salmon. PLoS ONE, 12: e0183781.
Wessel Ø., Hansen E.F., Dahle M.K., Alarcon M., Vatne N.A., Nyman I.B., et al. 2020. Piscine orthoreovirus-1 isolates differ in their ability to induce heart and skeletal muscle inflammation in Atlantic salmon (Salmo salar). Pathogens, 9: 1050.
Yang M., Cao S., Liu Y., Zhang Z., Zheng R., LI Y., et al. 2022. Performance verification of five commercial RT-qPCR diagnostic kits for SARS-CoV-2. Clinica Chimica Acta, 525: 46–53.
Zhang Y., Polinski M.P., Morrison P.R., Brauner C.J., Farrell A.A.P., Garver K.A. 2019. High-load reovirus infections do not imply physiological impairment in salmon. Frontiers in Physiology, 10: 114.
Zhao J., Vendramin N., Cuenca A., Polinski M., Hawley L.M., Garver K.A. 2021. Pan-Piscine orthoreovirus (PRV) detection using reverse transcription quantitative PCR. Pathogens, 10: 1548.
Supplementary material
Supplementary Material 1 (XLSX / 420 KB).
- Download
- 420.18 KB
Supplementary Material 2 (DOCX / 40.8 KB).
- Download
- 40.84 KB
Supplementary Material 3 (DOCX / 25.0 KB).
- Download
- 25.03 KB
Information & Authors
Information
Published In

FACETS
Volume 8 • January 2023
Pages: 1 - 9
Editor: S.J. Cooke
History
Received: 13 October 2022
Accepted: 17 March 2023
Version of record online: 22 May 2023
Copyright
© 2023 Authors Fuller and Siah and The Crown. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All data are within the manuscript and supplementary material.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: MPP, MH, JP
Data curation: MPP, CAH, AS, CF
Formal analysis: MPP, AS, CF
Funding acquisition: MPP, MH, JP
Investigation: MPP, CAH, AS
Methodology: MPP, CAH, AS, CF
Project administration: MPP, MH
Resources: CAH, AS, CF, MH
Supervision: MPP, AS, MH
Validation: CAH, AS, CF
Visualization: MPP, MH
Writing – original draft: MPP
Writing – review & editing: MPP, JP
Competing Interests
The authors have declared that no competing interests exist.
Funding Information
Funding for this project was provided by the Department of Fisheries and Oceans Canada through the Program for Aquaculture Regulatory Research (PARR) to MPP and MH.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Mark P. Polinski, Colleen A. Haddad, Ahmed Siah, Chad Fuller, Mark Higgins, and Jay Parsons. 2023. British Columbia freshwater salmon hatcheries demonstrate minimal contribution to piscine orthoreovirus (PRV) regional occurrence with no evidence for nonendemic strain introductions. FACETS.
8(): 1-9. https://doi.org/10.1139/facets-2022-0218
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item