Incorporating anthropogenic thresholds to improve understanding of cumulative effects on seagrass beds
Abstract
Cumulative human impact analysis is a promising management tool to estimate the impacts of stressors on ecosystems caused by multiple human activities. However, connecting cumulative impact scores to actual ecosystem change at appropriate spatial scales remains challenging. Here, we calculated cumulative effects (CE) scores for 187 seagrass beds in Atlantic Canada that accounts for both bay-scale and local-scale anthropogenic activities. We then developed a CE threshold to evaluate where degradation of seagrass beds from multiple human activities is more likely. Overall, the CE score was the best predictor of human impacts for seagrass beds. Locations with high watershed land alteration and nitrogen loading had the highest CE scores; however, we also identified seagrass beds with high CE scores in regions characterized by generally low levels of human activities. Forty-nine seagrass beds exceeded the CE threshold and, of these, 86% had CE scores that were influenced by three or more stressors that cumulatively amounted to a large score. This CE threshold approach can provide a simplified metric to identify areas where management of cumulative effects should be prioritized and further highlights the need to consider multiple human activities when assessing anthropogenic impacts to coastal habitats.
Introduction
Cumulative human impact (CHI) mapping has emerged as a promising management tool to better account for and estimate the impacts of stressors caused by multiple human activities in marine systems (Batista et al. 2014; Halpern et al. 2019). CHI mapping methods combine knowledge of the distribution and intensity of multiple human activities with estimates of the vulnerability of different habitats (or species) to individual stressors, into a single comparable cumulative effects (CE) score (Halpern et al. 2008). While CHI mapping cannot reveal possible synergistic or antagonistic effects among multiple stressors, this additive CE approach provides a simplified metric to evaluate risk to marine habitats that can more easily be communicated for management decisions.
Most CHI mapping in the marine realm has focused on broadscale quantification of human activities, having been applied globally (Halpern et al. 2008, 2019) or regionally (Selkoe et al. 2009; Micheli et al. 2013; Clarke Murray et al. 2015; Afflerbach et al. 2017). These maps are typically developed on a ≥1 km2 grid, making it difficult to assess CE for individual habitats that occur over smaller spatial scales. Further, those maps that do quantify CE in smaller grids (<1 km2) often include stressors that have been scaled down from global estimates, making these maps difficult to apply to local and regional management decisions (Holon et al. 2015). Broadscale CE maps are also less relevant for biogenic habitats, like seagrass or kelp beds, that are influenced by not only broadscale human activities (e.g., watershed land use) but also those occurring at finer, more local scales (e.g., aquaculture or shoreline alterations) (Shelton et al. 2017).
Improving impact predictions and providing insight into what areas represent priorities for coastal zone management can be achieved by incorporating thresholds into cumulative effects analyses (Halpern and Fujita 2013). However, this is challenging given the unknowns in thresholds for various anthropogenic activities, as well as the specificity of thresholds to habitat types, locations, and potentially other factors (e.g., season) (Wu et al. 2017). The identification of thresholds and the level of disturbance (i.e., quantity of pollutant, intensity of stressor) beyond which substantial ecosystem changes are expected, are common applications in environmental management to prevent dramatic shifts in ecosystem structure and functioning (Groffman et al. 2006). While discrete thresholds are useful in areas dominated by a single human activity, this is rarely the case in coastal habitats that are increasingly subjected to multiple co-occurring stressors (Lotze et al. 2006; Crain et al. 2008; Murphy et al. 2019). Empirically derived thresholds that can be used to predict ecosystem risk from cumulative impacts are not readily available given the lack of data on how multiple anthropogenic stressors combine and impact marine ecosystems (Griffiths et al. 2020; Stockbridge et al. 2020). Furthermore, managers are typically only presented with the past, present, and (or) future human activities occurring in an area, rather than the explicit levels of environmental stress resulting from those human activities. Thus, a different yet complementary estimation approach for CE thresholds is necessary.
Seagrass is a globally distributed marine plant that provides many valuable ecosystem services, including nursery habitat, sediment stabilization, and carbon storage (Barbier et al. 2011; Campagne et al. 2014; McDevitt-Irwin et al. 2016). Seagrass habitats provide an ideal case study to estimate a CE threshold and evaluate its use as the influence of human activities, and associated threshold effects have been well studied relative to other Canadian coastal marine habitats (Murphy et al. 2021). Human activities and related stressors have been linked to seagrass degradation and decline worldwide (Waycott et al. 2009; Short et al. 2011; Dunic et al. 2021), with the cumulative effect of localized stressors playing an important role (Telesca et al. 2015). Thus, increased attention to the CE of multiple human activities is a priority for improving management and conservation of seagrass ecosystems (Griffiths et al. 2020). In Canada, the dominant seagrass species, eelgrass (Zostera marina), creates important biogenic habitat on all three coasts, has been designated an ecologically significant species by Fisheries and Oceans Canada, and is identified as an important habitat to include in management and conservation planning (DFO 2009a; Rubidge et al. 2020; Murphy et al. 2021). While federal legislation relevant to seagrass protection may consider CE (Modernized Fisheries Act Bill C-68 section 34.1(1)(d)), there has been little focus on quantifying the magnitude of CE specific for seagrass habitats. Previous assessments have quantified the magnitude of a suite of human activities occurring in or near seagrass beds in Canada, yet none of these have combined the extent of human activities using a CE approach (Robb 2014; Iacarella et al. 2018; Murphy et al. 2019; van den Heuvel et al. 2019; Nahirnick et al. 2020; Nagel et al. 2021). Assessing human activities within seagrass beds and seagrass inhabited bays from a CE perspective would provide an overall representation of human impacts that accounts for differences in seagrass vulnerability to different activities and stressors.
Here, we calculate CE scores for 187 seagrass beds located along the coast of Atlantic Canada using a widely applied CHI methodology (Halpern et al. 2008). This work builds on an earlier study by Murphy et al. (2019), who quantified the intensity of multiple human impacts (five bay-scale and four local-scale) individually. While they provided a qualitative ranking of stressor intensity, they were not able to weight the relative importance of the human impacts according to seagrass vulnerability, preventing a more quantitative ranking of importance or an overall cumulative impact score. Now we aim to assess the combined impact of human activities near seagrass beds using a cumulative impact framework that translates the intensity of individual human activities into overall impact using knowledge of seagrass vulnerabilities. Applying this framework distills the cumulative effect of multiple stressors into a single estimate of relative impact on seagrass beds that is comparable across multiple scales. We also advance the application of the CHI methodology, by combining published thresholds for the individual stressors included in the calculation of our CE scores into a single CE threshold. We compare our CE scores against this threshold to evaluate how many seagrass beds may be at higher risk for degradation from multiple stressors and to identify locations where management of cumulative impacts should be prioritized. Finally, we discuss CE thresholds as a tool for impact assessments, and how CE scores and thresholds can strengthen conservation and management decisions compared to using scores for individual stressors alone.
Methods
Seagrass bed locations
We calculated CE scores for 187 eelgrass (Z. marina) beds, hereafter referred to as seagrass beds, located in 54 bays along the coast of Atlantic Canada. Seagrass bed locations were compiled from field surveys conducted over the past decade (Weldon et al. 2009; Schmidt et al. 2012; Skinner et al. 2013; Cullain et al. 2018; Wong 2018; Krumhansl et al. 2020). Of the 187 beds, 180 were included in Murphy et al. (2019), while seven additional seagrass bed locations were included from Krumhansl et al. (2021). The seagrass bed locations span three provinces (Nova Scotia (NS), New Brunswick (NB), and Prince Edward Island (PEI)) and two bioregions (Scotian Shelf and Gulf of St. Lawrence). Seagrass beds in these bioregions typically experience different oceanographic conditions, with a lower range in water temperature and a higher range of wave exposure on the Scotian Shelf coastline (DFO 2009b). However, seagrass beds in both regions are also subjected to short-term physical processes that are independent of regional processes, such as the daily heating and cooling of sheltered and shallow beds (Krumhansl et al. 2020).
Cumulative effects scores
To calculate a CE score for each seagrass bed, we applied an additive methodology (sensu Halpern et al. 2008) that quantifies cumulative impacts by combining the intensity of individual stressors with impact weights specific to eelgrass ecosystems, then sums these individual impacts into a single CE score. While human activities can also interact to influence ecosystems synergistically or antagonistically (Crain et al. 2008; Stockbridge et al. 2020), most CE studies assume additive impacts given the vast unknowns for other types of interactions (Korpinen and Andersen 2016; Hodgson et al. 2019). Cumulative impact scores for seagrass (IS) were thus calculated as:where n is the number of human activities, Di is the normalized value of intensity of activity i at each seagrass bed, Sj is the presence of seagrass ecosystems (and thus reduces to 1), and ui,j is the impact weighting (or habitat vulnerability) of seagrass bed j to human activity i (Halpern et al. 2008).
(1)
We used human activities as proxies for stressors (e.g., urban watershed land use is a proxy for pollution run-off) since in most cases direct measurements of stressors are not available for all seagrass beds in the region. Therefore, we refer to the intensity of human activities as stressor scores (i.e., the Di). Six stressors were chosen for their expected influence on coastal biogenic habitats like seagrass: urban watershed land use, agricultural watershed land use, nitrogen loading, invasion extent, near-field shellfish aquaculture activity, and hardened shoreline (Table 1). We did not include climate change related stressors as we chose to focus on stressors that could be managed on local or regional scales. To calculate stressor scores for urban watershed land use, agricultural watershed land use, and nitrogen loading, we used intensities from the standardized coastal human impact metric previously published in Murphy et al. (2019). While the three land-based activities included in the CE score (urban land use, agriculture land use, and nitrogen loading) are interrelated and have similar root causes, we include each of them separately in the CE score because they each represent different stressors to seagrass (urban runoff and inorganic pollution, sediment input, and nutrient input, respectively; Table 1) that can elicit different plant responses. While the quantity of inorganic nitrogen loading is a better predictor of seagrass coverage than total nitrogen loading (Coffin et al. 2021), we use estimates of total nitrogen loading taken from nitrogen loading models in the CE score since nitrate concentration data were not widely available across the 187 sites included in this study. We conducted a sensitivity analysis for selected sites where both measures were available and determined that using total-N loading to represent relative anthropogenic nutrient input instead of nitrate-N loading does not result in major changes to the overall CE scores (Supplementary Material A.2 and C).
Table 1.
| Human activity | Intensity measure | Corresponding stressor | Vulnerability score |
|---|---|---|---|
| Nutrient pollution | Human-derived nitrogen loading rate (kg N ha bay−1 year−1) | Nutrient input: into oligotrophic waters (for NS and NB sites) Nutrient input: into eutrophic waters (for PEI sites) | 2.3 2.9 |
| Hardened shoreline | Anthropogenically hardened shoreline, % | Coastal engineering: altered flow dynamics | 2.8 |
| Invasion extent | Number of invasive biofouling species considered established | Invasive species (from ballast, etc.) | 2.6 |
| Watershed land use—agriculture | Watershed agriculture land cover, % | Sediment input: increase | 2.2 |
| Shellfish aquaculture activity | Lease area within 25 m radius standardized to marine area | Aquaculture: shellfish | 2.0 |
| Watershed land use—urban | Watershed urban land cover, % | Average of two stressors related to urban land alteration: urban runoff and inorganic pollution | 1.95 |
Note: NS, Nova Scotia, NB, New Brunswick, PEI, Prince Edward Island.
We adjusted the calculation of two additional stressors in Murphy et al. (2019), invasion extent and near-field shellfish aquaculture, to more closely link these stressors with previously published thresholds (see below). Invasion extent was adjusted to represent the number of established invasive biofouling species (≥3 consecutive years of sightings) as opposed to a 10-year average of invader presence/absence. Murphy et al. (2019) included two sub-measures for shellfish aquaculture (near-field (<25 m) and mid-field (<1 km)) and only considered presence/absence of shellfish aquaculture within these ranges. Here, we measure shellfish aquaculture activity as the area of shellfish aquaculture leases within a 25 m radius surrounding each seagrass bed, standardized to the marine area within the 25 m radius. We excluded four activities included in Murphy et al. (2019) (overwater structures, water quality, riparian land alteration, and fishing activity) from the CE score as these activities do not have published thresholds available and it is currently unclear at what levels these activities become detrimental to seagrass. Finally, we included one additional stressor not previously included in Murphy et al. (2019) to represent the impact of hardened shorelines on seagrass beds (Patrick et al. 2014). We used Google Earth (Google Earth Pro, 2021) and the most recent and clearest imagery data available for each site (ranging between 2014 and 2021) to measure the percentage of shoreline that had been anthropogenically hardened (including riprap, bulkheads, bridge, and wharf abutments, etc.) within 1 km of each seagrass bed.
Vulnerability scores for seagrass habitats (i.e., the ui,j) have not been developed for Atlantic Canada; thus, we used a vulnerability matrix developed for the nearby New England region that includes vulnerability scores (also called impact weights) for 14 marine ecosystems, including seagrass (Kappel et al. 2012a, 2012b). Current evidence supports the application of vulnerability matrices developed in a specific area to different regions (Kappel et al. 2012a). These vulnerability scores were developed using a standardized survey design to gather knowledge from experts on criteria that are expected to influence the vulnerability of an ecosystem to a stressor (Halpern et al. 2007; Teck et al. 2010). The five vulnerability criteria considered by experts in the calculation of a single vulnerability score for each ecosystem–stressor combination included: (i) the spatial scale that a stressor will impact an ecosystem, (ii) the frequency of the stressor, (iii) the trophic impact and extent of marine life affected by a stressor within an ecosystem, (iv) the degree to which the ecosystem’s “natural” state is changed by the stressor, and (v) the time it will take for the affected components of the ecosystem to return to their former state following the impact (Kappel et al. 2012a). The vulnerability matrix developed by Kappel et al. (2012a) provides vulnerability scores for 58 stressors. We use the vulnerability scores specific to seagrass habitat from Kappel et al. (2012a) and link each of the six human activities included in the CE score to the dominant stressor expected to be caused by that human activity (Table 1). Nutrient pollution was further separated into region-specific stressors with separate vulnerability scores since the PEI bays and estuaries are generally considered eutrophic while those in NS and NB are generally considered oligotrophic.
Before weighting stressors based on vulnerability, the intensities of the six individual stressors were normalized (i.e., adjusted to range between 0 and 1), with zero representing the absence of the stressor and one representing the highest observed value within the region plus 10%. We added 10% to the maximum observed value for each impact in our dataset as this range has been suggested as appropriate to capture current and likely near-term future levels of the stressors and are often used in CE analyses (Halpern and Fujita 2013). An example of the CE score calculation can be found in Supplementary Material A, Table S1.
We assessed differences in CE scores among and within regions using 95% confidence intervals around the medians (Chambers et al. 1983). For regional comparisons, we split the Gulf of St. Lawrence into the mainland coast (Gulf NS + NB) and PEI. We grouped seagrass beds into five CE classes (very low to very high impact) based on the Jenks natural breaks classification method, which groups CE scores into classes by minimizing within-class variability and maximizing between-class variability. We conducted a Principal Components Analysis (PCA) using the six individual stressor scores and the CE score for each seagrass bed to identify the stressors that explained the most variation in the magnitude of human activities across seagrass beds. We centered and scaled variables before analysis. We considered any variable that had a loading that contributed more than one variable worth of information (>0.378) as an important contributor to the principal component. The PCA was conducted using the vegan package in R (Oksanen et al. 2021).
Cumulative effects threshold
The CE scores we calculated for all 187 seagrass beds are relative to each other and to the total number of stressors considered in the calculation. To relate the CE scores to actual impacts we would expect human activities to have on seagrass, we calculated a CE threshold score that was derived from existing threshold values for each individual stressor. We conducted a literature review to identify previously derived threshold levels for each of the six individual stressors. For some stressors, thresholds specific to seagrass habitat were not available so we used thresholds related to aquatic vegetation or coastal health in general. Table 2 outlines the individual threshold levels used in the calculation of the CE threshold score and the rationale behind the selection of each threshold. We used the thresholds for the individual stressors to estimate the CE threshold using the same methods and formula described above for the CE scores but replaced the intensity of each stressor with the identified threshold levels (Table 2; see Supplementary Material A, Table S2 for calculation). Therefore, the CE threshold score is the same across all 187 eelgrass beds and was estimated to be 3.17 in NS and NB sites, and 3.26 in PEI sites (i.e., IT = 3.17 or 3.26). We evaluated which seagrass bed CE scores exceeded this threshold (i.e., which IS > IT) to assess how many seagrass beds may be at risk for degradation from multiple stressors. Finally, we assessed how many of the six individual stressor scores exceeded the corresponding individual threshold levels for all 187 seagrass beds.
Table 2.
| Human activity | Intensity measure | Threshold | Rationale |
|---|---|---|---|
| Nutrient pollution | Human-derived nitrogen loading rate (kg N ha·bay−1·year−1) | 100 kg N·ha−1 year−1 | • Estimated 50% eelgrass decline when total nitrogen is approximately 100 kg N·ha−1·year−1 (Valiela et al. 1997) • Nitrogen loading shown to be detrimental to eelgrass coverage at loading rates of 50–100 kg N·ha−1·year−1 (Latimer and Rego 2010). • Inorganic nitrogen loads of 57 kg Nitrate-N·ha−1·year−1 has been linked to 50% decline in eelgrass cover (Coffin et al. 2021). |
| Hardened shoreline | Shoreline anthropogenically hardened, % | 5.4% | • Submerged aquatic vegetation abundance was lower in sub-estuaries with >5.4% hardened shoreline (i.e., riprap; Patrick et al. (2014). |
| Invasion extent | Number of invasive biofouling species established | 1 | • Established biofouling invaders can lead to shading, breakage, and reduced growth of aquatic vegetation (Wong and Vercaemer 2012, Carman and Grunden 2010, Moore et al. 2018). • Even low biofouling on eelgrass shoots can result in modest impacts to eelgrass growth (Long and Grosholz 2015). • Persistent species observance for 2–3 consecutive years generally considered established (C. DiBacco DFO Aquatic Invasive Species Group, personal communication, June 2020). |
| Watershed land use—agriculture | Watershed agriculture land cover, % | 19% | • See Supplementary Material A, Table S3 for literature search results |
| Shellfish aquaculture activity | Marine area covered by shellfish aquaculture leases within 25 m radius, % | 20% | • Oyster aquaculture leases exceeding 20% density spatially compete with eelgrass (Wagner et al. 2012). • Shellfish aquaculture leases within 25 m of seagrass beds can lead to shading and physical damage of seagrass (Vance 2014, Skinner et al. 2013). |
| Watershed land use—urban | Watershed urban land cover, % | 21.5% | • Greater than 20% watershed developed land is related to significant negative effects on submerged aquatic vegetation (Li et al. 2007). • See Supplementary Material A, Table S4 for literature search results |
Results
Cumulative effects scores
We observed a gradient of CE scores for seagrass beds in Atlantic Canada ranging from 0.14 to 7.05 (Fig. 1a). The maximum possible CE score (i.e., if each stressor was at the maximum observed value within a single site) was 11.65. However, this maximum score is unlikely given the improbability of all six stressors reaching maximum levels at a site. Relative to all 187 seagrass beds assessed, those along the coast of PEI (n = 66) had the highest CE scores (median = 3.77 ± 0.42 95% CI; Fig. 2). Median CE scores for seagrass beds in the Gulf NB + NS region (1.23 ± 0.18, n = 97) were also significantly lower than median CE scores in the Scotian Shelf region (2.02 ± 0.29, n = 24).
Fig. 1.
Fig. 2.
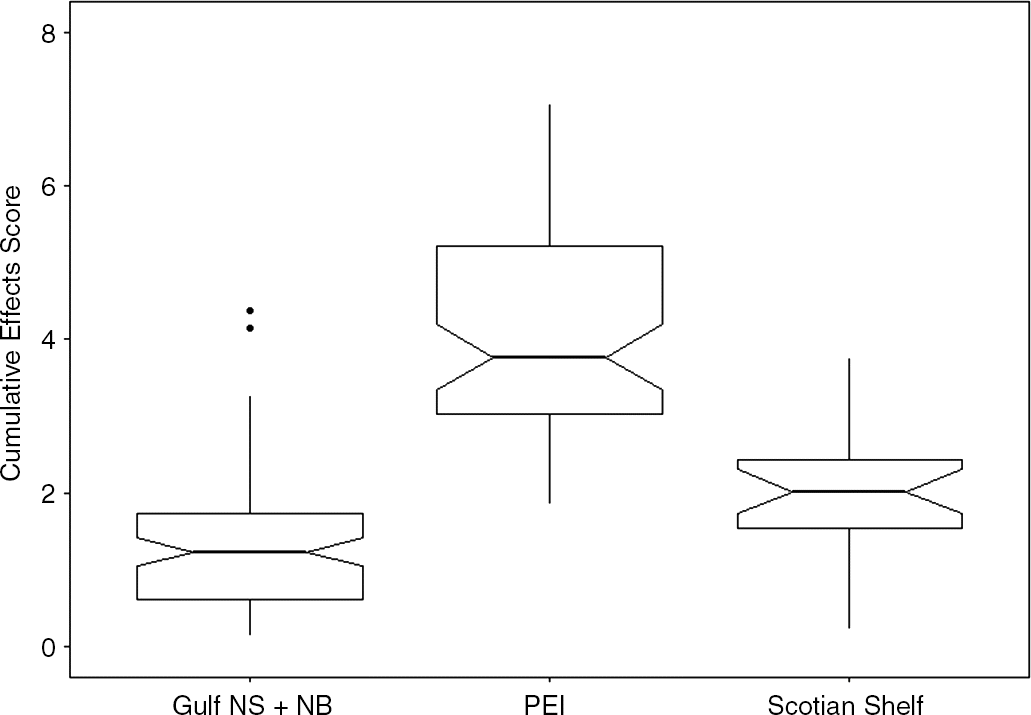
The lowest CE score across all 187 beds (0.14) was in the north bay of St. Simon in Gulf NB + NS. While the median CE score for Gulf NB + NS seagrass beds was also the lowest among the three regions, beds in this region experienced a wide range of CE classes, with eight beds classed as very high or high and 40 beds classed as very low. Scotian Shelf seagrass beds were generally classified as relatively low impact, with no seagrass beds in the Scotian Shelf region classed as very high; however, four were classed as high (three located in St. Margaret’s Bay and one located near Crescent Beach; Fig. 1a). The Scotian Shelf region also had three seagrass beds with the second lowest CE scores among all 187 beds (0.23), all located along the Eastern Shore of Nova Scotia. Twenty-nine seagrass beds in the Gulf region were classed as having very high CE scores; all but one of these beds were in PEI.
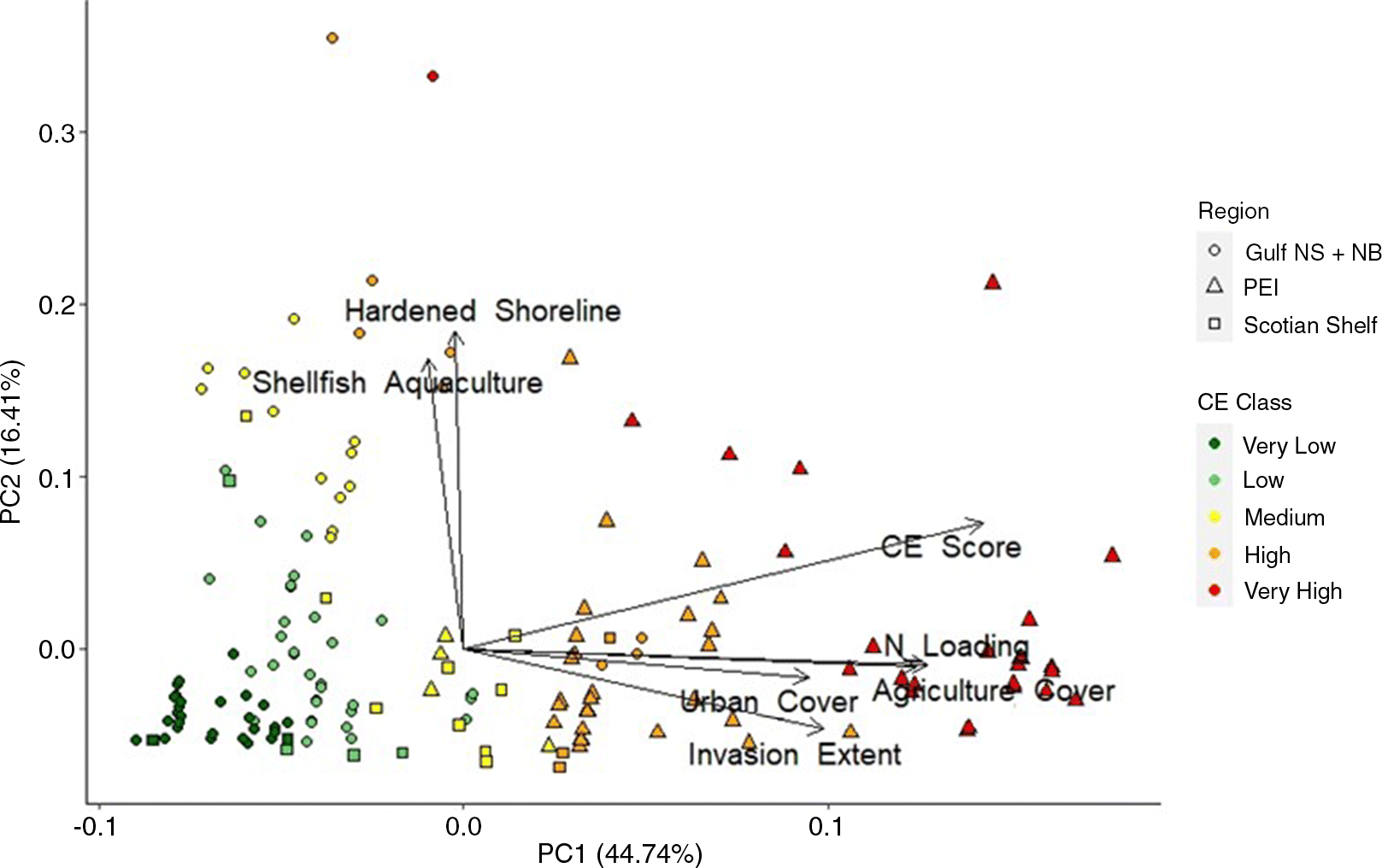
The first principal component (PC1) in the PCA of the CE scores and individual stressor scores explained 45% of the variation across the seagrass beds (Fig. 3). CE scores were the best predictors of separation among groups along PC1, with other large positive loadings for agriculture cover and nitrogen loading (Fig. 3; Supplementary Material A, Table S6). The second (PC2) and third (PC3) principal components explained 16% and 14% of the variance across seagrass beds, respectively. Together, these axes captured a gradient in hardened shorelines and shellfish aquaculture activity among seagrass bed locations. Both stressors had large positive loadings for PC2 and were mainly associated with beds in Gulf NB + NS and PEI. On PC3, hardened shorelines had large positive loadings and shellfish aquaculture had large negative loadings, and distinguished seagrass beds with near-field shellfish aquaculture from those without (Supplementary Material A, Table S6).
Fig. 3.
Assessing the risk of degradation from multiple stressors using a cumulative effects threshold
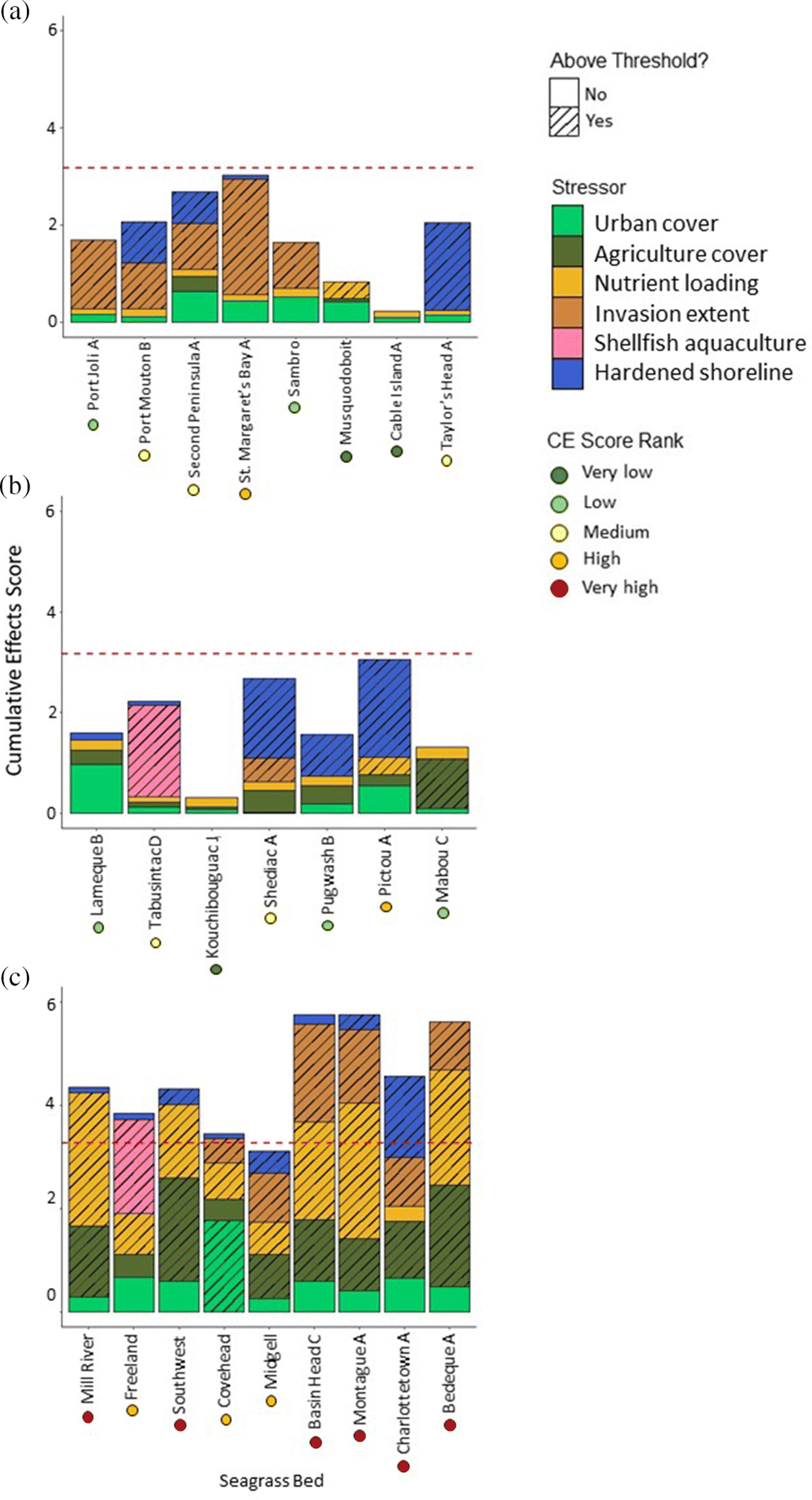
Forty-nine of the 187 seagrass beds (26%) had CE scores that exceeded the CE threshold (Fig. 4). Of these, the majority (88%) were in PEI (Fig. 1b). Of the 66 seagrass beds in PEI, 65% were above the CE threshold (Fig. 1b). Only five (10%) seagrass beds in Gulf NS + NB and one (2%) Scotian Shelf seagrass bed were above the CE threshold (Fig. 1b). Seventeen seagrass beds had CE scores within 10% of the threshold, indicating they are at risk of exceeding the threshold if nearby human activities were to increase. This includes three seagrass beds in the Scotian Shelf, four seagrass beds in Gulf NB + NS, and 10 seagrass beds in PEI (Supplementary Material B).
Fig. 4.
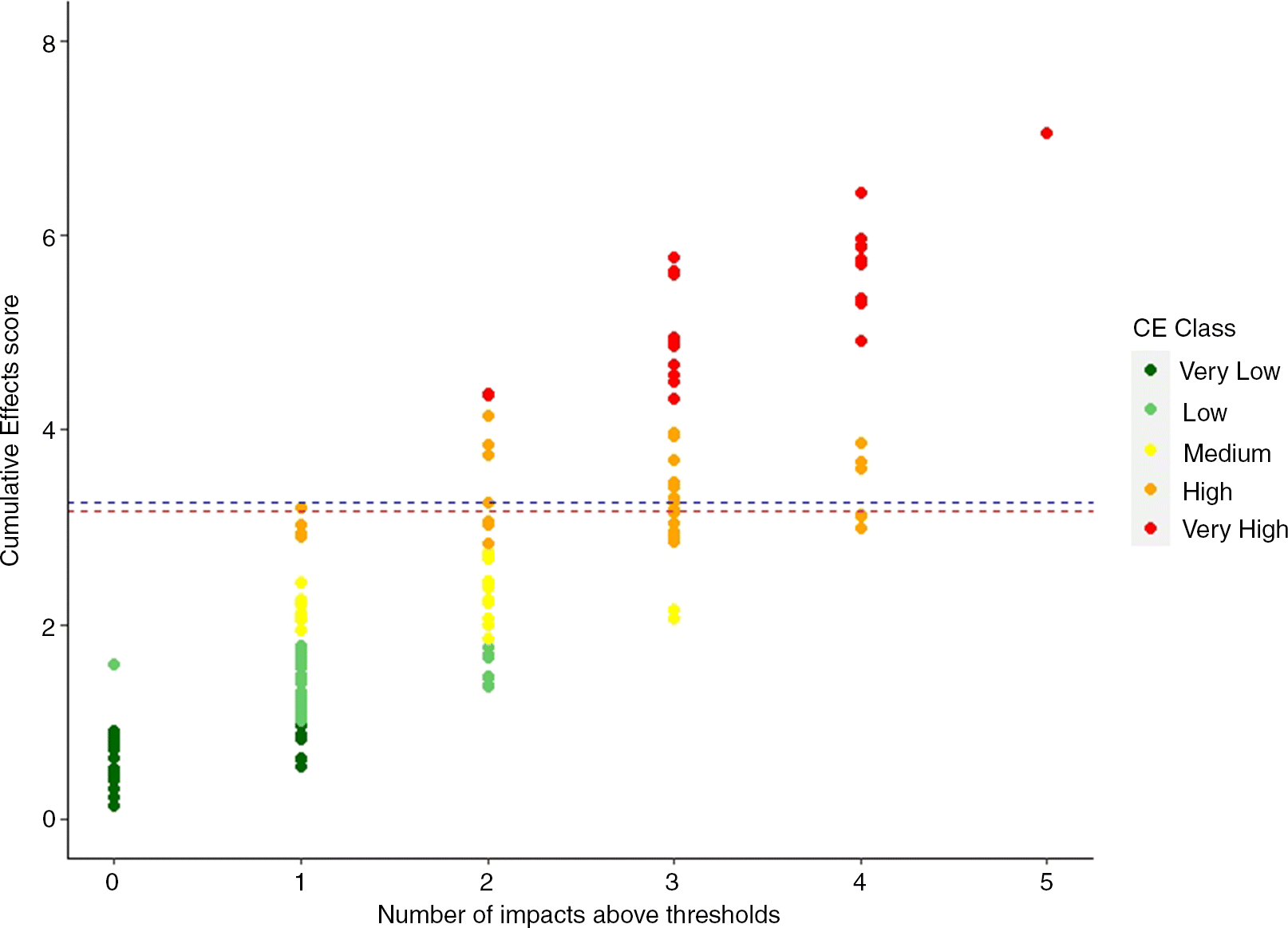
Seagrass beds that exceeded the CE threshold typically had multiple stressors that exceeded their individual thresholds (Fig. 4). Thirty-six seagrass beds did not exceed any of the individual stressor thresholds and, as expected, most CE scores of these beds were classified as very low. Fifty-nine seagrass beds had only one individual stressor score exceed the corresponding threshold. Most of these beds had CE scores ranging from very low to low; however, some of the CE scores were classed as high and one exceeded the CE threshold (Cocagne.H). The majority (79%) of seagrass beds where two component stressors exceeded their individual thresholds still had CE scores below the estimated CE threshold (Fig. 4). Only six seagrass beds had two stressors that exceeded their individual thresholds and a CE score that also exceeded the estimated CE threshold (Supplementary Material A, Fig. S5). In contrast, the majority of seagrass beds where three, four, or five component stressors exceeded their individual threshold levels also had CE scores that exceeded the estimated CE threshold (62%, 76%, and 100%, respectively). Not surprisingly, in many cases the seagrass beds identified in Murphy et al. (2019) as highly impacted also had among the highest CE scores (e.g., Pictou and Mill River in the Gulf of St. Lawrence, and St. Margaret’s Bay in the Scotian Shelf; Fig. 5). Similarly, low-impacted seagrass beds identified in Murphy et al (2019) also typically had the lowest CE scores (e.g., Kouchibouguac in the Gulf of St. Lawrence and Cable Island in the Scotian Shelf; Fig. 5).
Fig. 5.
Discussion
Applying a CE approach to a recent analysis of human activities near seagrass beds in Atlantic Canada has revealed several insights regarding the influence of multiple stressors on seagrass beds. First, CE scores were the best predictors of human impacts among seagrass beds across a broad spatial scale. Next, vulnerability weights used in the calculation of CE scores provided a more holistic assessment of human impact near seagrass beds compared to individual assessments of stressors (i.e., Murphy et al. 2019). Finally, CE threshold score estimates can provide a simplified metric to evaluate risk to marine habitats that is easily communicated for management applications, with subsequent repercussions for conservation decisions.
Our results revealed three distinct gradients of human activity across the 187 seagrass beds. The first, and strongest, gradient was defined by variation in CE scores. Seagrass beds with high CE scores were more often located in bays with high human-derived nitrogen loading and high watershed land alteration, and to a lesser extent, the presence of invasive species and high urban watershed land alteration. While nutrient loading and land alteration were the largest contributors to the overall CE score, this is not because they had the highest vulnerability scores and were given more weight in the CE score calculation. In fact, two activities (invasion extent and hardened shorelines) had higher vulnerability scores (Table 1) and were therefore considered more detrimental to seagrass ecosystem health and, in turn, weighted more heavily in the CE score. Instead, our results show that seagrass beds are more likely influenced by the combination of human activities that cause multiple stressors that contribute to high CE scores. For example, of the 71 seagrass beds with high agricultural land cover within the watershed (>19%), 87% also had high impact scores (i.e., above defined thresholds from Table 2) for at least two other human activities and 25% had high impact scores for at least three other human activities. Seagrass and other near-shore ecosystems have experienced the fastest increase in cumulative human impacts in marine systems globally, and intensification of land-based pressures has contributed to this fast pace of change (Halpern et al. 2019). The stressors resulting from land-use intensification are often considered to be among the most significant threats to seagrass ecosystems (Quiros et al. 2017; Lefcheck et al. 2018; van den Heuvel et al. 2019); therefore, accounting for the full array of stressors resulting from land use, including sediment and pollution run-off, nutrient loading, wave energy alteration, and others, is important to accurately assess CE to seagrass and other nearshore ecosystems.
Our results also revealed a second gradient of human activities defined by variation in shellfish aquaculture and hardened shoreline. Higher CE scores were found in seagrass beds with a greater proportion of the adjacent shoreline anthropogenically hardened (i.e., riprap, breakwater, etc.) that also had higher coverage of shellfish aquaculture leases within 25 m. In our calculation of CE scores for seagrass beds in Atlantic Canada we included a combination of bay-scale and local-scale human activities. Management strategies that reduce specific local-scale stressors have been shown to enhance the resilience of coastal habitats to global stressors (Strain et al. 2015). Therefore, cumulative human impact maps that identify local-scale stressors, like adjacent hardened shoreline or nearfield shellfish aquaculture, and the cumulative effect of these stressors, can be an important management tool. However, fine-scale cumulative human impact mapping across an entire region is difficult and most fine-scale mapping efforts are limited to single bays (but see Holon et al. 2015), making coast-wide comparisons difficult. Using a CE approach to combine bay-scale and local-scale human activities for a single habitat of interest across an entire region, as we have done here, can support management and conservation planning by providing detailed information on local-scale CE rather than scaling down from broader cumulative human impact maps that would not be as precise.
We expected high CE scores for PEI seagrass beds given the high intensity of nutrient loading and shellfish aquaculture activity in the province, and many of these seagrass beds were located in closer proximity to centers of high human populations than for seagrass beds in other provinces. However, our results also revealed high CE scores in some seagrass beds in the Scotian Shelf and Gulf NS + NB regions that were less expected given the relatively lower influence of human activities in those regions. For example, two Scotian Shelf seagrass beds (Crescent Beach and St. Margaret’s Bay A) had high CE scores, with Crescent Beach exceeding the CE threshold and St. Margaret’s Bay A with a CE score within 5% of the CE threshold. The high scores in these two beds were driven by a combination of high established invasive species richness and either another activity that exceeded the individual threshold (i.e., hardened shoreline for Crescent Beach) or several activities that did not exceed their individual thresholds but cumulatively amounted to a higher CE score (i.e., St Margaret’s Bay A). The ability to identify anomalous locations highly impacted by human activities within a region that is otherwise characterized by generally low CE scores is an important aspect of the CE approach and would allow targeted management actions. These results also highlight the importance of considering multiple marine and land-based stressors when assessing human impacts to coastal ecosystems. While there are other seagrass beds in the Scotian Shelf region with high established invasive species richness (e.g., Port l’Hebert with 4 established invasive species), our results show that the impact of cumulative human activities is of less concern at these sites given their low CE scores. This knowledge is particularly important for informing restoration and management plans, as focusing on the impacts of only one dominant stressor (e.g., invasive species) may obscure the complete picture of anthropogenic influence on a site (Batista et al. 2014).
CE analyses typically include several related human activities, given that multiple stressors with different mechanisms of action on ecosystem components and with different levels of intensity, can result from similar activities (Knights et al. 2013; Clarke Murray et al. 2014). In our study, we included three interrelated land-based impacts in the CE score as they represented different stressors to seagrass (urban runoff and inorganic pollution, sediment input, and nutrient input) that could elicit different plant responses. While agricultural land cover and nutrient loading were correlated (r = 0.71), the nutrient loading metric incorporates inputs from several other sources including atmospheric deposition, wastewater, and industrial outputs. Since the nutrient loading stressor score already accounts for fertilizer run-off from agricultural activities, we used agricultural land cover as a proxy for sediment input as we did not have direct measurements of this stressor. Including both nutrient loading and sediment input in CE scores for seagrass is important since excess nutrient loading primarily impacts seagrass through reduced light attenuation from algae overgrowth, while excess sediments can have negative impacts through burial of shoots (de Boer 2007) or by changing sediment chemistry (Krause-Jensen et al. 2011). Despite having similar root causes, the importance of nutrient loading and sediment input (agricultural cover) in the CE scores supports the inclusion of multiple stressor pathways with different mechanisms of action to provide a more holistic picture of the complex network of impacts important for seagrass health.
Cumulative effects thresholds as a tool for seagrass management and conservation
Our development of the CE threshold approach translates the (relative) CE score into a potential measure of habitat (or ecosystem) status that can be used to identify areas where degradation from multiple human activities is more likely. Of the 49 seagrass beds that exceeded the CE threshold, 86% had CE scores that were highly influenced by three or more stressors that cumulatively amounted to a large score. This emphasizes the need to consider multiple human activities when assessing anthropogenic impact to seagrass (or any coastal) habitat. Considering activities in a singular manner will fail to capture the cumulative stress on seagrass beds that are subjected to multiple human activities. Only six seagrass beds that exceeded the CE threshold had only two dominant human activities and only one seagrass bed with CE scores exceeding the CE threshold had only one dominant human activity (Supplementary Material A, Fig. S5). However, in the majority of these seagrass beds, the CE score would not have exceeded the CE threshold if only the one or two dominant human activities were considered. Each of these seagrass beds were also influenced by other human activities that did not exceed the individual stressor thresholds, with these activities contributing cumulatively to exceed the CE threshold.
Effective recovery and restoration of seagrass habitat requires careful site selection and management approaches that limit the impacts caused by multiple human activities (van Katwijk et al. 2016, Grech et al. 2017). Given that the tolerance of marine ecosystems to stressors will vary, linking CE scores to CE thresholds allows identification of ecosystems that will be negatively impacted by multiple human activities. Consideration of CE thresholds can also aid in marine protected area (MPA) planning by providing knowledge of the risk of degradation from multiple activities in surrounding areas, which can affect the success of an MPA or other spatial conservation measure (e.g., marine park, marine refuge/reserve). Often the limited funds available for restoration or conservation initiatives means that plans need to prioritize the best economic and ecological returns; using CE thresholds to identify those areas may be beneficial.
The CE threshold we have calculated is a conservative estimate, as we only included activities or stressors for which we could find published quantitative thresholds. Other human activities and stressors that are known to affect seagrass beds, such as boating activity (e.g., Unsworth et al. 2017) or overwater structures (e.g., Burdick and Short 1999), could also be important, but were not included in our analysis since thresholds were difficult to identify. Thresholds for seagrass growth have been identified for several environmental factors and their interactions (e.g., light, temperature; Lee et al. 2007; Adams et al. 2020), yet further work to link the magnitude of human activities to environmental thresholds is necessary to accurately predict how seagrass beds will respond to CE of human activities, including synergistic or antagonistic effects (Crain et al. 2008). The thresholds for human activities we have included in the CE threshold estimate could also be improved in the future by linking them more explicitly to environmental thresholds known to influence seagrass (i.e., measuring the direct effects of a stressor rather than using a proxy). Furthermore, approaches that could link the magnitude of human activities to environmental thresholds would be a valuable tool, increasing our ability to make predictions for how seagrass would be expected to respond to the CE of human activities.
Caveats and next steps
There are a number of caveats to the calculation of CE scores and thresholds we present here, along with further research that should be done in the future to expand and improve upon this work. First, the vulnerability scores used were not specific to Atlantic Canada and instead were previously developed for the New England region (Kappel et al. 2012a). Comparisons of the results of the same elicitation approach applied to separate groups of experts in California vs. New England supports the generalizability of applying vulnerability weights across regions (Kappel et al. 2012a); however, differences in environmental conditions between or within regions may alter the response of seagrass to stressors (Krumhansl et al. 2021). For example, warmer temperatures increase the light requirements of seagrass and can consequently increase their vulnerability to stressors that limit underwater light (Beca-Carretero et al. 2018). Ongoing work by Fisheries and Oceans Canada to revise the vulnerability scores from Kappel et al. (2012a, 2012b) to better reflect Atlantic Canadian marine habitats may also provide additional insight on inter-regional differences in vulnerability of coastal habitats to human activities and advance the CE scores for this region in the future.
We calculated the CE threshold scores using the same thresholds for all seagrass beds; therefore, the score only varied slightly due to our use of differing nitrogen loading vulnerability weights for PEI versus NS and NB beds. However, the static CE threshold scores (i.e., IT = 3.17 and 3.26) may actually vary considerably depending on the environmental context. For example, comprehensive nutrient loading thresholds developed by Coffin et al. (2021) for PEI seagrass beds are useful to assess the risk of seagrass decline from nutrient loading in a region dominated by agricultural land use. However, these thresholds are not applicable to seagrass in NS and NB given different oceanographic conditions experienced by seagrass beds between the two regions (i.e., micro- vs. macro-tidal estuarine conditions), and the use of inorganic nitrogen load data used to develop the empirical relationship from which the threshold response was estimated. While total nitrogen load (the values available for our inter-regional comparison), would be similar to inorganic nutrient load in areas dominated by agriculture (like PEI), the composition would vary in NS and NB depending on the main source of nitrogen. These inter-regional differences in data availability made it difficult to apply thresholds specific for inorganic nitrogen across the 187 seagrass beds we assess in this paper. Further work to include pertinent environmental information to adapt thresholds across different regions could increase the suitability of the thresholds used and, in turn, the accuracy of the CE threshold score. In addition, since seagrass is more influenced by nitrate-N loading rate than total-N loading rate (van den Heuvel et al. 2019), the CE scores could be refined in the future by using dissolved inorganic nitrogen concentration data to represent anthropogenic nutrient loading, should such information become available for all seagrass sites. Given that nitrate concentration data are not always readily available, the use of nitrogen loading models to estimate total nitrogen loading rates provides a more widely applicable method to estimate CE and aligns with a more precautionary approach to identify impacted seagrass beds, throughout Atlantic Canada (as the CE thresholds were lower when using total nitrogen versus nitrate loading rates; see Supplementary Material A.2).
The information available for each human activity (and its associated stressors and mechanism of action) varies greatly, presenting more of a data gap rather than a knowledge-based one for cumulative impact analyses. While land-based stressors (nutrient loading, sediment input) appeared to be dominant components of the CE score for many seagrass beds, this may be an artefact of having less accurate intensity estimates for lesser-known stressors (i.e., invasive species extent vs. nutrient loading), or having excluded other types of marine-based stressors altogether (e.g., overwater structures, fishing activity, etc.). Human stressors additional to those included in our CE score occur in the region and may influence seagrass ecosystems, yet could not be incorporated owing to incomplete data availability or estimated threshold levels for seagrass. For example, these include coastal commercial fishing activity, boating activity, and overwater structures as presented in Murphy et al. (2019), but may also include historical stressors that have legacy impacts (i.e., pollutants from coastal gold mining operations) or stressors whose impacts are only beginning to be acknowledged (i.e., lost or discarded fishing gear). Further research to quantify the extent to which these activities impact seagrass habitat in the region and the appropriate threshold levels past which seagrass degradation is likely would allow inclusion into the CE score and threshold calculations, improving the accuracy of this assessment.
Most importantly, future work should validate the CE scores presented here using seagrass field data to evaluate how the CE scores correlate with seagrass ecosystem conditions. Species distribution models could also be used for validation to assess whether the CE scores we have calculated here can help explain areas of the coast where seagrass beds could exist based on environmental conditions (i.e., temperature, substrate type) but do not, perhaps because of the impacts from multiple human activities. Cumulative impact assessments are rarely validated using empirical data and those that have been done have reported conflicting relationships. Andersen et al. (2015) found that cumulative human impact measures were negatively related to biodiversity status, whereas other studies have observed only weak relationships between estimated cumulative impacts and other indicators of ecosystem condition such as benthic community abundance and composition (Clark et al. 2016). Specifically for seagrass, Stockbridge et al. (2021) found that cumulative impact assessments were not a good indicator of seagrass bed structure (e.g., percent cover, shoot density, etc.). Given that few validation studies have been conducted, and those that have been done typically use very different indicators of ecosystem condition, further work to validate cumulative impact assessments is necessary. Additionally, the vulnerability weights used in the calculation of CE scores were assigned by expert opinion; therefore, they should also be validated with empirical data (Bevilacqua et al. 2018). Finally, the CE model used in this paper assumes that stressors combine additively, which is highly unlikely in real-world scenarios (Crain et al. 2008). Multi-factorial laboratory and field experiments to assess interactions between stressors and inform whether stressors in cumulative impact assessments should combine additively, synergistically, or antagonistically would improve the accuracy of CE scores. Whether the cumulative impact assessment and associated vulnerability weights and CE threshold we applied in this paper are good indicators of ecosystem condition for Atlantic Canadian seagrass beds is currently unclear. Formally linking CE scores and thresholds to seagrass condition will provide insight into the usefulness of our CE assessment approach for seagrass ecosystems, and identify areas for improvement. Future research to validate how cumulative impact assessments translate to real-world seagrass ecosystem condition in Atlantic Canada will be crucial for the optimal management of these important coastal ecosystems.
Acknowledgements
We thank our colleagues Cathryn Clarke Murray and Kira Krumhansl (Fisheries and Oceans Canada) for helpful discussions, as well as two anonymous reviewers whose constructive comments improved the manuscript.
Competing interests statement
The authors declare there are no competing interests.
References
Adams MP, Koh EJY, Vilas MP, Collier CJ, Lambert VM, Sisson SA, et al. 2020. Predicting seagrass decline due to cumulative stressors. Environmental Modelling and Software, 130: 104717.
Afflerbach JC, Yocum D, and Halpern BS. 2017. Cumulative human impacts in the Bering Strait region. Ecosystem Health and Sustainability, 3: 1379888.
Andersen JH, Halpern BS, Korpinen S, Murray C, and Reker J. 2015. Baltic Sea biodiversity status vs. cumulative human pressures. Estuarine, Coastal and Shelf Science, 161: 88–92.
Barbier E, Hacker S, Kennedy C, Koch E, Stier A, and Silliman B. 2011. The value of estuarine and coastal ecosystem services. Ecological Monographs, 81: 169–193.
Batista MI, Henriques S, Pais MP, and Cabral HN. 2014. Assessment of cumulative human pressures on a coastal area: Integrating information for MPA planning and management. Ocean and Coastal Management, 102: 248–257.
Beca-Carretero P, Olesen B, Marbà N, and Krause-Jensen D. 2018. Response to experimental warming in northern eelgrass populations: comparison across a range of temperature adaptations. Marine Ecology Progress Series, 589: 59–72.
Bevilacqua S, Guarnieri G, Farella G, Terlizzi A, and Fraschetti S. 2018. A regional assessment of cumulative impact mapping on Mediterranean coralligenous outcrops. Scientific Reports. 8: 1757.
Burdick DM, and Short FT. 1999. The effects of boat docks on eelgrass beds in coastal waters of Massachusetts. Environmental Management, 23: 231–240.
Campagne CS, Salles JM, Boissery P, and Deter J. 2014. The seagrass Posidonia oceanica: Ecosystem services identification and economic evaluation of goods and benefits. Marine Pollution Bulletin, 97: 391–400.
Carman MR, and Grunden DW. 2010. First occurrence of the invasive tunicate Didemnum vexillum in eelgrass habitat. Aquatic Invasions, 5: 23–29.
Chambers JM, Cleveland WS, Kleiner B, and Tukey PA. 1983. Comparing Data Distributions. In Graphical Methods for Data Analysis. pp. 47–74.
Clarke Murray C., Mach ME., and Martone, RG. 2014. Cumulative effects in marine ecosystems: scientific perspectives on its challenges and solutions. WWF-Canada and Center for Ocean Solutions. 60 pp. Available from awsassets.wwf.ca/downloads/cumulativeeffects__updated_forwebupload_singlepages.pdf
Clarke Murray C, Agbayani S, Alidina HM, and Ban NC. 2015. Advancing marine cumulative effects mapping: An update in Canada’s Pacific waters. Marine Policy, 58: 71–77.
Clark D, Goodwin E, Sinner J, Ellis J, and Singh G. 2016. Validation and limitations of a cumulative impact model for an estuary. Ocean and Coastal Management, 120: 88–98.
Coffin MRS, Poirier LA, Clements JC, Dickson E, Guyondet T, Crane CJ, and van den Heuvel MP. 2021. Dissolved oxygen as a Marine Environmental Quality (MEQ) measure in upper estuaries of the southern Gulf of St. Lawrence: Implications for nutrient management and eelgrass (Zostera marina) coverage. Canadian Science Advisory Secretariat Research Document. 2021/056. Iv + 27 p.
Crain CM, Kroeker K, and Halpern BS. 2008 Interactive and cumulative effects of multiple human stressors in marine systems. Ecology Letters, 11: 1304–1315.
Cullain N, McIver R, Schmidt AL, and Lotze HK. 2018. Spatial variation of macroinfaunal communities associated with Zostera marina beds across three biogeographic regions in Atlantic Canada. Estuaries and Coasts, 41: 1381–1396.
De Boer WF. 2007. Seagrass-sediment interactions, positive feedbacks and critical thresholds for occurrence: a review. Hydrobiolgia, 591: 5–24.
DFO. 2009a. Does eelgrass (Zostera marina) meet the criteria as an ecologically significant species? DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2009/018.
DFO. 2009b. Development of a framework and principles for the biogeographic classification of Canadian marine areas. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2009/056.
Dunic JC, Brown CJ, Connolly RM, Turschwell MP, and Cote IM. 2021. Long-term declines and recovery of meadow area across the world’s seagrass bioregions. Global Change Biology 00: 1–14.
ESRI. 2021. World ocean basemap. [online]: Available from arcgis.com/home/item.html?id=67ab7f7c535c4687b6518e6d2343e8a2
Grech A, Hanert E, McKenzie L, Rasheed M, Thomas C, Tol S, et al. 2017. Predicting the cumulative effect of multiple disturbances on seagrass connectivity. Global Change Biology, 24: 3093–3104.
Griffiths LL, Connolly RM., and Brown CJ. 2020. Critical gaps in seagrass protection reveal the need to address multiple pressures and cumulative impacts. Ocean and Coastal Management, 183.
Groffman PM, Baron JS, Blett T, Gold AJ, Goodman I, Gunderson LH, et al. 2006. Ecological thresholds: the key to successful environmental management or an important concept with no practical application?. Ecosystems, 9: 1–13.
Halpern BS, Selkoe KA, Michel F, and Kappel CV. 2007. Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conservation Biology, 21: 1301–1315.
Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D’Agrosa C, et al. 2008. A global map of human impact on marine ecosystems. Science, 319: 948–952.
Halpern BS, and Fujita R. 2013. Assumptions, challenges, and future directions in cumulative impact analysis. Ecosphere, 4: art131–11.
Halpern BS, Frazier M, Afflerbach J, Lowndes JS, Micheli F, O’Hara C, et al. 2019. Recent pace of change in human impact on the world’s ocean. Scientific Reports, 9: 11609.
Hodgson EE, Halpern BS, and Essington TE. 2019. Moving beyond silos in cumulative effects assessment. Frontiers in Ecology and Evolution, 7: 211.
Holon F, Mouquet N, Boissery P, and Bouchoucha M. 2015. Fine-Scale Cartography of human impacts along French Mediterranean coasts: A relevant map for the management of marine ecosystems, 10: e0135473.
Iacarella JC, Adamczyk E, Bowen D, Chalifour L, Eger A, Heath W, et al. 2018. Anthropogenic disturbance homogenizes seagrass fish communities. Global Change Biology, 24: 1904–1918.
Kappel CV, Halpern BS, Selkoe KA, and Cooke RM. 2012a. Eliciting expert knowledge of ecosystem vulnerability to human stressors to support comprehensive ocean management. In Expert Knowledge and Its Application in Landscape Ecology. Edited by Perera A., Drew C., Johnson C. Springer, New York, NY.
Kappel CV, Halpern BS, and Napoli, N. 2012b. Mapping Cumulative Impacts of Human Activities on Marine Ecosystems (03.NCEAS.12). SeaPlan, Boston.
Knights AM., Koss RS, and Robinson LA. 2013. Identifying common pressure pathways from a complex network of human activities to support ecosystem-based management. Ecological applications, 23: 755–765.
Korpinen S, and Andersen JH. 2016. A global review of cumulative pressure and impact assessments in marine environments. Frontiers in Marine Science, 3: 153–164.
Krause-Jensen D., Carstensen J., Nielsen SL, Dalsgaard T, Christensen PB, Fossing H, et al. 2011. Sea bottom characteristics effect depth limits of eelgrass Zostera marina. Marine Ecology Progress Series 425: 91–102.
Krumhansl K, Dowd M, Wong MC. 2020. A characterization of the physical environment at seagrass sites along the Atlantic coast of Nova Scotia. Canadian Technical Report of Fisheries and Aquatic Sciences 3361. Fisheries and Oceans Canada. iv + 213 p.
Krumhansl KA, Dowd M, and Wong MC. 2021. Multiple metrics of temperature, light, and water motion drive gradients in eelgrass productivity and resilience. Frontiers in Marine Science, 8: 597707.
Latimer JS, and Rego SA. 2010. Empirical relationship between eelgrass extent and predicted water- shed-derived nitrogen loading for shallow New England estuaries. Estuarine, Coastal and Shelf Science, 90: 231–240.
Lee K-S, Park SR, and Kim YK. 2007. Effects of irradiance, temperature, and nutrients on growth dynamics of seagrasses: A review. Journal of Experimental Marine Biology and Ecology, 350: 144–175.
Lefcheck JS, Orth RJ, Dennison WC, Wilcox DJ, Murphy RR, Keisman J, et al. 2018. Long-term nutrient reductions lead to the unprecedented recovery of a temperate coastal region. Proceedings of the National Academy of Sciences of the United States of America, 115: 3658–3662.
Li X, Weller DE, Gallegos CL, Jordan TE, and Kim HC. 2007. Effects of watershed and estuarine characteristics on the abundance of submerged aquatic vegetation in Chesapeake Bay Subestuaries. Estuaries and Coasts, 30: 840–854.
Long HA, and Grosholz ED. 2015. Overgrowth of eelgrass by the invasive colonial tunicate Didemnum vexillum: Consequences for tunicate and eelgrass growth and epifauna abundance. Journal of Experimental Marine Biology and Ecology, 473: 188–194.
Lotze, HK, Lenihan, HS, Bourque, BJ, Bradbury RH, Cooke, RG, Kay, MC, et al. 2006. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science, 312: 1806–1809.
McDevitt-Irwin JM, Iacarella JC, Baum JK. 2016. Reassessing the nursery role of seagrass habitats from temperate to tropical regions: a meta-analysis. Marine Ecology Progress Series, 557: 133–143.
Micheli F, Halpern BS, Walbridge S, Ciriaco S, Ferretti F, Fraschetti S, et al. 2013. Cumulative human impacts on Mediterranean and black sea marine ecosystems: assessing current pressures and opportunities. PLoS ONE, 8: e79889.
Moore AM, Lowen JB, and DiBacco C. 2018. Assessing invasion risk of Didemnum vexillum to Atlantic Canada. Management of Biological Invasions, 9: 11–25.
Murphy GEP, Wong MC, and Lotze HK. 2019. A human impact metric for coastal ecosystems with application to seagrass beds in Atlantic Canada. FACETS 4: 210–237.
Murphy GEP, Dunic JC, Adamczyk EM., Bittick SJ, Côté IM, Cristiani J, et al. 2021. From coast to coast to coast: ecology and management of seagrass ecosystems across Canada. FACETS, 6: 139–179.
Nagel EJ, Murphy G, Fast J, Bittick SJ, Adamczyk EM, O’Connor MI, et al. 2021. Application of a coastal human impact metric and nitrogen loading model to ten eelgrass (Zostera marina) meadows in British Columbia. Canadian Technical Report of Fisheries and Aquatic Sciences, 3429: vii + 45 p.
Nahirnick NK, Costa M, Schroeder S, and Sharma T. 2020. Long-term eelgrass habitat change and associated human impacts on the West coast of Canada. Journal of Coastal Research, 36: 30–40.
Oksanen J, Simpson GL, Blanchet FG, Kindt R, Legendre P, and Minchin PR. 2021. vegan: Community Ecology Package. R package version 2.6-2. [online]: Available from CRAN.R-project.org/package=vegan
Patrick CJ, Weller DE, Li X, and Ryder M. 2014. Effects of shoreline alteration and other stressors on submerged aquatic vegetation in subestuaries of Chesapeake Bay and the mid-Atlantic coastal bays. Estuaries and Coasts, 37: 1516–1531.
Quiros TEA, Croll D, Tershy C, Fortes MD, and Raimondi P. 2017. Land use is a better predictor of tropical seagrass condition than marine protection. Biological Conservation, 209: 454–463.
Robb CK. 2014. Assessing the impact of human activities on British Columbia’s estuaries. PLoS ONE, 9: e99578.
Rubidge E, Jeffery S, Gregr EJ, Gale, KSP, and Frid, A. 2020. Assessment of nearshore features in the Northern Shelf Bioregion against criteria for determining Ecologically and Biologically Significant Areas (EBSAs). Canadian Science Advisory Secretariat Research Document. 2020/023: vii + 63.
Schmidt AL, Wysmyk JKC, Craig SE, and Lotze HK. 2012. Regional-scale effects of eutrophication on ecosystem structure and services of seagrass beds. Limnology and Oceanography, 57: 1389–1402.
Selkoe KA, Halpern BS, Ebert CM, Franklin EC, Selig ER, Casey KS, et al. 2009. A map of human impacts to a “pristine” coral reef ecosystem, the Papahānaumokuākea Marine National Monument. Coral Reefs, 28: 635–650.
Shelton AO, Francis TB, Feist BE, Williams GD, Lindquist A, and Levin PS. 2017. Forty years of seagrass population stability and resilience in an urbanizing estuary. Journal of Ecology, 105: 458–470.
Short, FT, Polidoro, B, Livingstone, SR, Carpenter, KE, Bandeira, S, Bujang, JS, et al. 2011. Extinction risk assessment of the world’s seagrass species. Biological Conservation, 144: 1961–1971.
Skinner MA, Courtenay SC, and McKindsey CW. 2013. Reductions in distribution, photosynthesis, and productivity of eelgrass Zostera marina associated with oyster Crassostrea virginica aquaculture. Marine Ecology Progress Series, 486: 105–119.
Stockbridge J, Jones AR, and Gillanders BM. 2020. A meta-analysis of multiple stressors on seagrasses in the context of marine spatial cumulative impacts assessment. Scientific Reports, 10: 1–11.
Stockbridge J, Jones AR, Gaylard SG, Nelson MJ, and Gillanders BM. 2021. Evaluation of a popular spatial cumulative impact assessment method for marine systems: A seagrass case study. Science of the Total Environment, 780: 146401.
Strain EMA, van Belzen J, van Dalen J, Bouma TJ, and Airoldi L. 2015. Management of local stressors can improve the resilience of marine canopy algae to global stressors. PLoS ONE, 10: e0120837.
Teck SJ, Halpern BS, Kappel CV, Micheli F, Selkoe KA, Crain CM, et al. 2020. Using expert judgment to estimate marine ecosystem vulnerability in the California Current. Ecological Applications, 20: 1402–1416.
Telesca L, Belluscio A, Criscoli A, Ardizzone G, Apostolaki ET, Fraschetti S, et al. 2015. Seagrass meadows (Posidonia oceanica) distribution and trajectories of change. Science Reports, 5: 12505.
Unsworth RKF, Williams B, Jones BL, and Cullen-Unsworth LC. 2017. Rocking the boat: Damage to eelgrass by swinging boat moorings. Frontiers in Plant Science, 8: 1–11.
Valiela I, McClelland J, Hauxwell J, Behr PJ, Hersh D, and Foreman K. 1997. Macroalgal blooms in shallow estuaries: controls and ecophysiological and ecosystem consequences. Limnology and Oceanography, 42: 1105–1118.
van den Heuvel MR, Hitchcock JK, Coffin MRS, Pater CC, and Courtenary SC. 2019. Inorganic nitrogen has a dominant impact on estuarine eelgrass distribution in the Southern Gulf of St. Lawrence, Canada. Limnology and Oceanography, 64: 2313–2327.
van Katwijk MM, Thorhaug A, Marbà N, Orth RJ, Duarte CM, Kendrick GA, et al. 2016. Global analysis of seagrass restoration: The importance of large-scale planting. Journal of Applied Ecology, 53: 567–578.
Vance A. 2014. Applying an ecosystem-based risk management approach to the relationship between eelgrass beds and oyster aquaculture at multiple spatial scales in eastern New Brunswick, Atlantic Canada. Master of Marine Management thesis, Dalhousie University, Halifax, Nova Scotia. p. 34–37.
Wagner E, Dumbauld BR, Hacker SD, Trimble AC, Wisehart LM, and Ruesink JL. 2012. Density-dependent effects of an introduced oyster, Crassostrea gigas, on a native intertidal seagrass, Zostera marina. Marine Ecology Progress Series, 468: 149–160.
Waycott, M, Duarte, CM, Carruthers, TJB, Orth, RJ, Dennison, WC, Olyarnik, S, et al. 2009. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proceedings of the National Academy of Sciences, 106: 12377–12381.
Weldon J, Courtenay S, and Garbary D. 2009. The community aquatic monitoring program (CAMP) for measuring marine environmental health in coastal waters of the southern Gulf of St. Lawrence: 2007 overview. Canadian Technical Report of Fisheries and Aquatic Sciences 2825. viii + 75 p.
Wong MC. 2018. Secondary production of macrobenthic communities in seagrass (Zostera marina, eelgrass) beds and bare soft sediments across differing environmental conditions in Atlantic Canada. Estuaries and Coasts, 41: 536–548.
Wong MC, and Vercaemer B. 2012. Effects of invasive colonial tunicates and a native sponge on the growth, survival, and light attenuation of eelgrass (Zostera marina). Aquatic Invasions, 7: 315–326.
Wu PP, Mengersen K, McMahon K, Kendrick GA, Chartrand K, York PH, et al. 2017. Timing anthropogenic stressors to mitigate their impact on marine ecosystem resilience. Nature Communications, 8: 1263.
Supplementary material
Supplementary Material A
- Download
- 473.06 KB
Supplementary Material B
- Download
- 38.49 KB
Supplementary Material C
- Download
- 8.39 KB
Information & Authors
Information
Published In

FACETS
Volume 7 • Number 1 • January 2022
Pages: 966 - 987
Editor: Irene Gregory-Eaves
History
Received: 27 August 2021
Accepted: 5 May 2022
Version of record online: 7 July 2022
Copyright
© 2022 Her Majesty the Queen in Right of Canada and H. Lotze. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper and in the Supplementary Material.
Key Words
Sections
Subjects
Plain Language Summary
Determining thresholds of combined human impacts to seagrass in Atlantic Canada
Authors
Author Contributions
GEPM and NEK conceived and designed the study.
GEPM and NEK analyzed and interpreted the data.
All drafted or revised the manuscript.
Funding Information
Funding was provided by Fisheries and Oceans Canada.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Grace E.P. Murphy, Noreen E. Kelly, Heike K. Lotze, and Melisa C. Wong. 2022. Incorporating anthropogenic thresholds to improve understanding of cumulative effects on seagrass beds. FACETS.
7(): 966-987. https://doi.org/10.1139/facets-2021-0130
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item