Temporal diversity patterns for fungi and mites associated with decaying Sporobolus pumilus (Spartina patens) in the Minas Basin, Nova Scotia
Abstract
Coastal salt marshes provide many ecosystem services; however, little is known of the biology of decomposer taxa in these systems. This study employed aboveground and belowground litterbags to characterize the fungal and mite communities associated with the dominant salt marsh grass, Sporobolus pumilus, in the Minas Basin, Nova Scotia. Decomposition rates of aboveground and belowground tissues and environmental variables were quantified to contextualize temporal patterns in community composition. Aboveground litterbag mass loss peaked in July and decreased consistently over succeeding months, which positively correlated with fungal richness. Fungal and mite richness displayed inverse relationships over time, with mites gradually increasing in diversity before peaking in November, suggesting the presence of a complex detrital network where mites and fungi respond to different and possibly unrelated environmental cues. This study offers a first look at temporal community dynamics of two neglected groups of decomposers associated with S. pumilus in Canada.
Introduction
Litter decomposition and its implications for carbon storage and nutrient cycling is governed by taxonomically and functionally diverse decomposer communities. This occurs through a combination of litter composition and quality (Prescott et al. 2000; Yan et al. 2018), the metabolic ability of decomposers (López-Mondéjar et al. 2018), and species interactions between decomposers (Hättenschwiler et al. 2005), all of which may directly affect plant diversity and community composition (Mazzoleni et al. 2007). Soil and litter communities are mainly composed of bacterial and fungal decomposers as well as consumers that feed on plant detritus and fungal biomass (Berg and Bengtsson 2007). Fungal conditioning of litter increases its palatability to shredding invertebrates (Suberkropp and Arsuffi 1984; Danger et al. 2012), facilitating litter fragmentation. Further, fungal hyphae and spores are palatable to grazing taxa such as Collembola and Oribatid mites (Klironomos and Kendrick 1996; Pollierer and Scheu 2021) whose feeding behaviors structure fungal communities and enhance fungal activity (Lussenhop 1992).
Despite the importance of decomposer communities to ecosystem function, large gaps remain in the understanding of spatiotemporal structuring in many ecosystems (Guerra et al. 2020) such as North American salt marshes. Salt marshes are highly productive ecosystems that support a wide range of ecosystem services such as providing nursery habitat for fish and shellfish (Boesch and Turner 1984; Deegan et al. 2000), birds (Darnell and Smith 2004; Hughes 2004), carbon storage (Chmura et al. 2003; Alongi 2020), storm buffering (Shepard et al. 2011; Möller et al. 2014), and sediment transport into adjacent coastal ecosystems (Krest et al. 2000). Studies of fungal and bacterial decomposers in North American salt marshes have mainly focused on plants inhabiting the low marsh (Benner et al. 1984; Newell et al. 1989; Lyons et al. 2005), and mite communities of North American salt marshes have not yet been the subject of any published scientific study except for a brief mention of Oribatid mite diversity from a salt marsh in Cape Breton Highlands National Park in Nova Scotia (Behan-Pelletier et al. 1987). More research is needed to further characterize the community composition of salt marsh decomposer communities, particularly in the high marsh.
This study employed a litterbag experiment to characterize the poorly known diversity of fungi and mites associated with decomposing leaf litter of Sporobolus pumilus (Roth) P.M. Peterson & Saarela (formerly Spartina patens). Taxonomic data were also related to decomposition rates and environmental variables. We hypothesized that (i) aboveground plant material will decompose more quickly than belowground material, as root material is relatively recalcitrant and the frequently flooded sediments are oxygen-poor (Howes and Teal 1994); (ii) fungal richness and mite diversity will be positively correlated with decomposition rate; and (iii) mite richness will be positively correlated with fungal richness as mites are some of the most abundant fungus feeders in terrestrial systems.
Methods
Study locations
This study was conducted in the Wolfville salt marsh in Wolfville, Nova Scotia, Canada, on the megatidal Minas Basin, which experiences diurnal tides. The marsh faces northward onto the Cornwallis River, with a second fragment on the north bank. Three sites were chosen, with two situated in the western and eastern parts of the southern portion of the marsh, divided through the middle by Wolfville Harbour. The third site is located in the northern portion of the marsh, separated from the other two sites by the Cornwallis River (Fig. 1). Site 1 (45.094672°, −64.355659°) was situated in the southeast of the marsh. The site was lower than the other two, permitting more frequent tidal inundation. Wetter sediments allowed for greater cover by Sporobolus alterniflorus (Loisel.) P.M. Peterson & Saarela (formerly Spartina alterniflora), although the site is still dominated by Sporobolus pumilus. Nevertheless, the relative abundance of S. alterniflorus compared with sites 2 and 3 is the most obvious distinguishing feature among the three sites. Site 2 (45.097542°, −64.364201°) was in the southwest of the marsh, close to the bank of a creek, 0.75 km northwest of site 1. This site was higher than site 1 resulting in drier sediments and a virtual monoculture of S. pumilus. Plant cover was very dense at this site, lacking any bare sediment. Site 3 (45.104801, −64.365388°) was in the northern part of the marsh, 0.85 km north of site 2 and 1.35 km northwest of site 1, only a few meters away from the northern bank of the Cornwallis River, which was rapidly eroding the marsh edge. This site was also composed entirely of S. pumilus; however, plant cover was thinner than at site 2, with prominent patches of bare sediment between the plants. Individual plants tended to grow shorter here than at sites 1 and 2. Additional discussion of the marsh plant composition of the Wolfville salt marsh can be found in Chapman (1937), dating back to a period when the marsh was highly disturbed due to agricultural activity, where the author notes S. pumilus (as Spartina patens) formed isolated clumps rather than dominating the high marsh as it does now, except for an area in the northern portion of the marsh where S. pumilus was not collected for hay, and in this portion the grass was the dominant species.
Fig. 1.
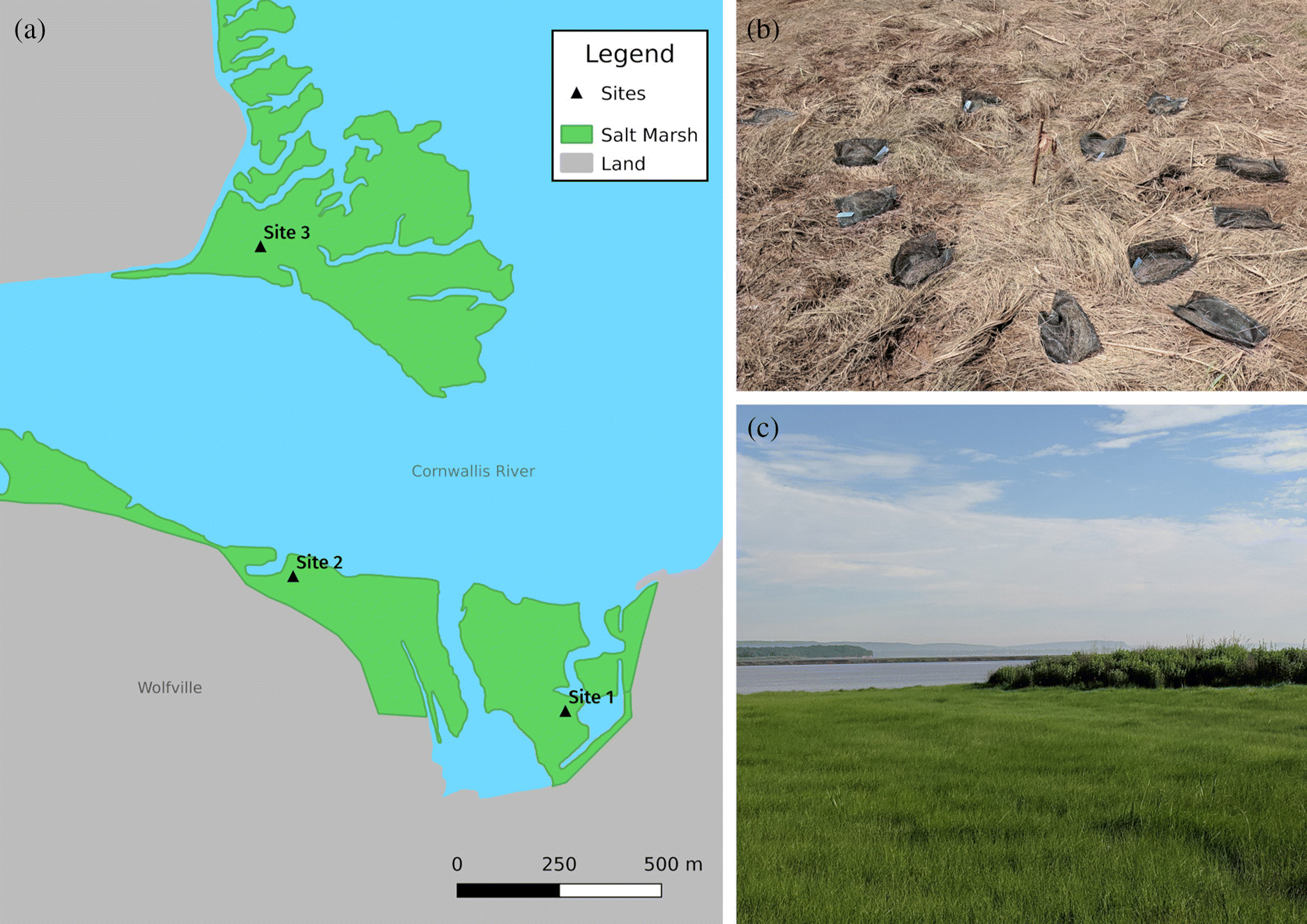
Litterbag design and retrieval
Overwintered shoot and root material of S. pumilus were collected from each of the three study sites on 15 May 2019. Due to the density of the clay-rich salt marsh sediments, roots could not be separated from the sediment in the field, so blocks of sediment were shoveled out of the marsh and transferred to the lab in large plastic bags. Extensive and careful washing was required to dissolve the sediments without breaking the fine root tips. Shoots and leaves were also washed to remove any attached sediments. Material was left to air dry at room temperature for three weeks to avoid killing the fungi already present in the grass.
The litterbags were made to be approximately 18 × 25 cm from nylon window screen with a 1.8 mm gap size and sealed with aluminum staples. Each bag was filled with 5.0 g of either shoot/leaf litter or dried roots derived from the specific site the litterbag was to be deployed in. Thirty-six bags were made for aboveground and belowground material for each of the three sites, in addition to some spares to account for potential loss of litter bags (244 litterbags in total). The litterbags were set up at the sites in early June 2019. At each site, 3 rebar poles were arranged in a triangle, each 5 m apart. The litterbags were tied to the poles with nylon fishing line, and bags containing roots were buried approximately 10 cm below the surface within a 2 m radius of the poles. Bags containing shoots and leaves were fastened to the ground with nails at the same distance as the belowground bags. Litterbags were retrieved at an interval of approximately 30 days, with the first retrieval in mid-June 2019, where one aboveground bag and one belowground bag was retrieved from each pole at each site, in addition to an extra aboveground and belowground bag from each site, totaling 24 retrieved bags per month. This initial retrieval, approximately 10 days after the litterbags were deployed, was to act as a control to determine how much mass loss occurred as a result of the process of preparing the bags, deploying them in the marsh, and returning them to the lab before significant decomposition had occurred.
Teabag decomposition
Decomposition was also measured using the tea bag index (Keuskamp et al. 2013). This method uses Lipton® (Unilever, Toronto Ontario) Green and Rooibos tea prepackaged in nonbiodegradable mesh bags as the substrate for decomposition, which makes this an easy, standardized method for measuring decomposition rates. The tea was first weighed including the bag and tag. Three bags of each kind of tea were buried 10 cm below the sediment surface at each site (18 bags in total). After 90 days, the bags were retrieved, the tea was removed from the bags and reweighed. Decomposition could then be quantified using the materials provided by teatime4science.org.
Environmental variables
At each collection interval, environmental variables were recorded for each site. Surface temperature and salinity were recorded monthly starting in July, soil moisture was recorded in October and November. Three measurements of each inorganic variable were recorded for each site. Surface temperature was recorded with an infrared thermometer, soil moisture was recorded with a Decagon (Meter Group, Pullman, Washington, United States) GS3 VWC moisture probe, and salinity was measured with an Oakton (Environmental Express, Charleston, South Carolina, United States) SALT 6+ handheld salinity meter.
Litterbag preparation for quantification of decomposition rates and diversity
After each litterbag retrieval, 18 of 24 retrieved litterbags were opened and the litter was washed free of sediment, air dried for one week, and weighed to determine mass loss (final weight compared to the original weight recorded during litterbag preparation). The remaining 6 litterbags (one aboveground and one belowground bag from each site) were used for quantifying the fungal richness and mite diversity. First, mites were extracted from the litterbag contents using 40 W Tullgren-Berlese funnels. See Krantz and Walter (2009) for detailed discussion of this sampling method. Following this, the litter was analyzed by microscopy to compile a list of reproductively active fungi present in the material. Belowground litterbags were found to host very few mites or reproductively active fungi, and so only aboveground bags were used for compiling diversity data.
Microscopy and specimen identification
The litter used for quantifying fungal biodiversity was carefully scanned under a dissecting microscope for fungal reproductive structures. Identification of species was done using light microscopy. Material was mounted predominantly in Windex (SC Johnson, Racine, Wisconsin, United States) (dilute ammonium hydroxide solution), as it is especially good at rehydrating dried fungal material, and Melzer’s solution, Lugol’s solution, and 5% potassium hydroxide were used for testing amyloidity or for the presence of ionomidotic reactions. Pretreatment with Congo Red or phloxine was used for staining nonrefractive material, where the former stains chitinous cell walls and the latter stains cytoplasm. Specimens were examined using a Nikon (Nikon Corporation, Tokyo, Japan) SMZ-2 T stereo microscope and a Leitz (Leica Microsystems GmbH, Wetzlar, Germany) SM-Lux compound microscope. A large body of taxonomic literature was used for identification, but the most important references for the identification to genus level were Sutton (1980); Ainsworth et al. (1973); Barr (2004); Barr (1990); Seifert et al. (2011); Kohlmeyer and Kohlmeyer (1979), and Jones et al. (2009). Generic monographs were used for species-level identification such as Shoemaker and Babcock (1989), as well as many type descriptions located in global species databases such as Index Fungorum (www.indexfungorum.org). Vouchers are deposited in the E.C. Smith herbarium at Acadia University, Nova Scotia, Canada, under the accession numbers ACAD21151F – ACAD21223F.
The mites collected in this study were stored in 70% ethanol until examined microscopically. Tullgren-Berlese extractions result in a large mix of mites of different species and instars which need to be sorted to record the richness and abundance of species present. One or two individuals of each morphospecies were first cleared in 85% lactic acid for 5–7 days, which macerates soft tissue, increasing light transmission through otherwise opaque material. Chitinous material is preserved such as the mite cuticle and fungal spores present in the gut, which has the added benefit of allowing for the observation of gut contents of fungivorous mite species. Cleared specimens were then mounted in polyvinyl alcohol (Bioquip Products Inc, Rancho Dominguez, California, United States) and dried on a heating mat at 40 °C for one week. Polyvinyl alcohol is a pre-prepared, water-soluble, semipermanent mountant with an ideal refraction index for light microscopy. Slide-mounted specimens were then identified using light microscopy primarily by consulting Gilyarov (1975, 1977, 1978); Balogh (1972); Dindal (1990); Krantz and Walter (2009); and Walter et al. (2014). Recent monographs were used when available, such as the monograph of Tydeidae by Silva et al. (2016), as well as national and international species checklists such as Behan-Pelletier and Lindo (2019); Subías et al. (2012); and Mąkol and Wohltmann (2012). The majority of mite species in North America are undescribed (Young et al. 2019), in most cases only allowing for identification to the genus level. Following identification of slide-mounted material, the remaining unsorted mites from each Tullgren-Berlese extraction were counted and sorted, with each morphospecies stored in 1.7 mL microcentrifuge tubes of 70% ethanol. Vouchers are deposited at the New Brunswick Museum in Saint John, New Brunswick, Canada.
Statistics
Data analysis and visualization were computed with the R software environment (R Core Team 2021), using the packages vegan (Oksanen et al. 2020), AICcmodavg (Mazerolle 2020), and ggplot2 (Wickham 2016). Vegan is a statistical package for community ecology, which was used for computing canonical correspondence analysis plots, an ordination technique used for relating community data to environmental variables. The technique is distinguished from other ordination methods in that it assumes that species respond normally along environmental gradients (ter Braak 1986).
The Akaike Information Criterion (AIC) is a model selection equation for determining which model best explains the variance in a dataset by weighing the goodness of fit of a model with the number of parameters included in it, where a larger number of parameters results in a greater penalty for the model due to the risk of overfitting. Expanding on this, the corrected Akaike Information Criterion (AICc) also incorporates sample size into its judgement of the models’ predictive value, making it useful for model selection with biological data (Anderson et al. 1998).
Results
Litterbag decomposition
Decomposition of litterbag contents (detrital Sporobolus grass) occurred most rapidly for aboveground litterbags containing shoots and leaves (p < 0.001) (Fig. 2a). Shoot-containing bags lost 1.72 ± 0.16 g (44% of initial mass) between June and November, and root-containing bags lost 1.02 ± 0.18 g (32% of initial mass). Between setup and the first collection interval, shoot-containing bags lost 0.19 ± 0.08 g (SE), while root-containing bags lost 0.57 ± 0.05 g (Fig. 2b). The initial interval between setup and June retrieval was intended to test how much mass would be lost due to handling rather than decomposition. Decomposition rate was not significantly different among the three sites for either root- or shoot-containing bags. Mass loss rates peaked in June and July before stabilizing by October and November. Soil temperature was positively correlated with shoot decomposition rate, whereas variation in root decomposition rate was best explained by a model involving salinity, soil moisture, fungal richness, and mite diversity (Table 1).
Fig. 2.
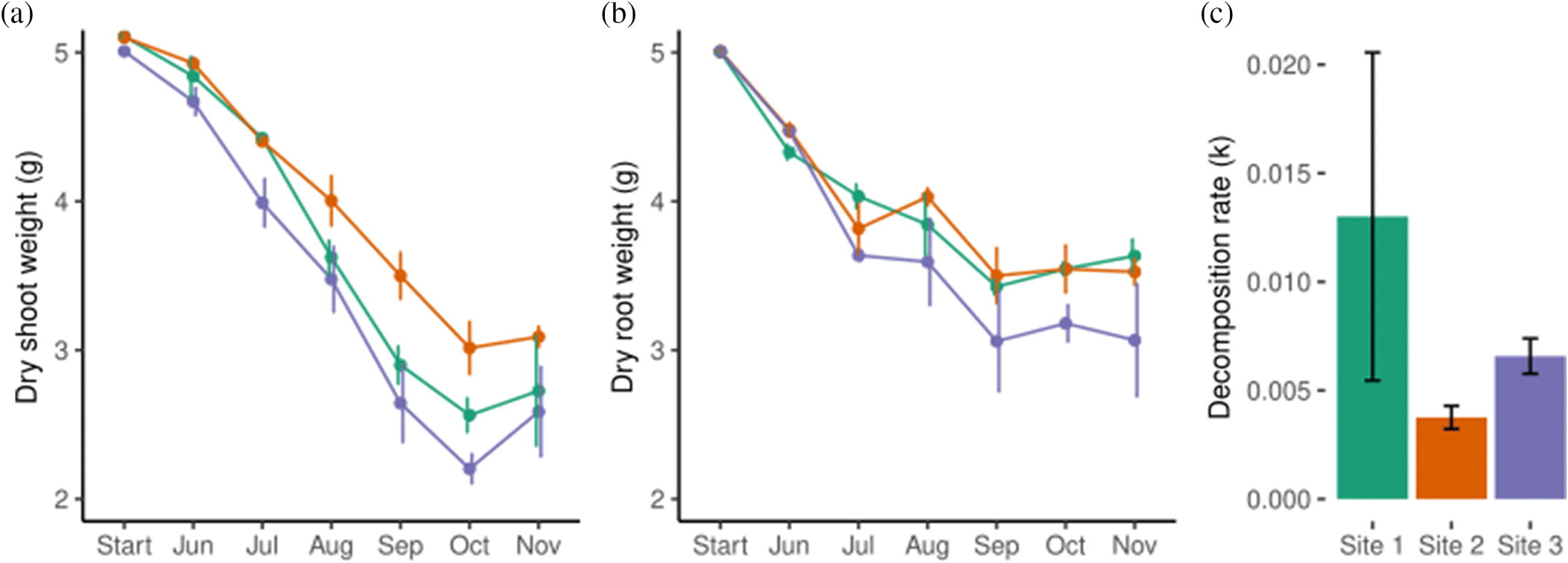
Table 1.
| Response | Predictor | B | Std. Error | t | R2 | df | F | p |
|---|---|---|---|---|---|---|---|---|
| Fungal richness | Intercept | −3.94 | 2.06 | −1.92 | — | — | — | 0.08 |
| Surface temperature | 0.07 | 0.09 | 0.77 | — | — | — | 0.5 | |
| Shoot litterbag weight | 1.85 | 0.76 | 2.42 | — | — | — | 0.03* | |
| Model summary | — | — | — | 0.49 | 2,11 | 7.28 | 0.009** | |
| Mite diversity | Intercept | 2.19 | 0.49 | 4.50 | — | — | — | 0.0006*** |
| Shoot litterbag weight | −0.35 | 0.15 | −2.42 | — | — | — | 0.03* | |
| Model summary | — | — | — | 0.26 | 1,13 | 5.85 | 0.03* | |
| Shoot decomposition rate | Intercept | −0.13 | 0.19 | −0.69 | — | — | — | 0.5 |
| Surface temperature | 0.04 | 0.01 | 0.01 | — | — | — | 0.01* | |
| Model summary | — | — | — | 0.39 | 1,12 | 9.29 | 0.01* | |
| Root decomposition rate | Intercept | 2.94 | 0.78 | 3.78 | — | — | — | 0.004** |
| Salinity | 0.02 | 0.005 | 4.29 | — | — | — | 0.002** | |
| Soil moisture | −0.04 | 0.009 | −4.22 | — | — | — | 0.002** | |
| Fungal richness | 0.09 | 0.02 | 3.69 | — | — | — | 0.004** | |
| Mite diversity | −0.29 | 0.11 | −2.67 | — | — | — | 0.02* | |
| Model summary | — | — | — | 0.77 | 4,10 | 12.73 | 0.0006*** |
Note:
Each model achieved the lowest AICc (corrected Akaike Information Criterion) given the following combinations of variables: biotic response variables used surface temperature, salinity, shoot litterbag weight, and soil moisture as predictors. Decomposition rates used surface temperature, salinity, soil moisture, fungal richness, mite diversity as predictors. * denotes p < 0.05, ** denotes p < 0.01, and *** denotes p < 0.001.
Of the environmental variables recorded from the three study sites, surface temperature and soil salinity did not differ significantly between sites. However, while moisture was not significantly lower at site 3 compared to sites 1 and 2 overall (F(2,15) = 2.7, p = 0.1), it should be noted soil moisture data were only collected for the months of October and November. Along with the observed difference in topology and plant community composition, sediments at sites 1 and 2 were probably somewhat wetter than site 3 (Supplementary Table S1).
Tea bag decomposition assay
Fourteen of 16 tea bags were recovered from the sites; two at site 1 were lost. Decomposition rate constant (k) was significantly higher at site 3 than site 2, indicating a higher mass loss rate (Fig. 2c). One of the tea bags at site 1 was much lighter than all the others in the study and this resulted in high standard error. The stabilization factor S represents the proportion of litter that is resistant to decomposition. S value was significantly lower at site 3 than at site 2, and again standard error was very high at site 1. 80.41% ± 8.27 of the rooibos tea dry weight remained after 90 days, and 31.05% ± 3.51 of the green tea remained.
Fungal community composition
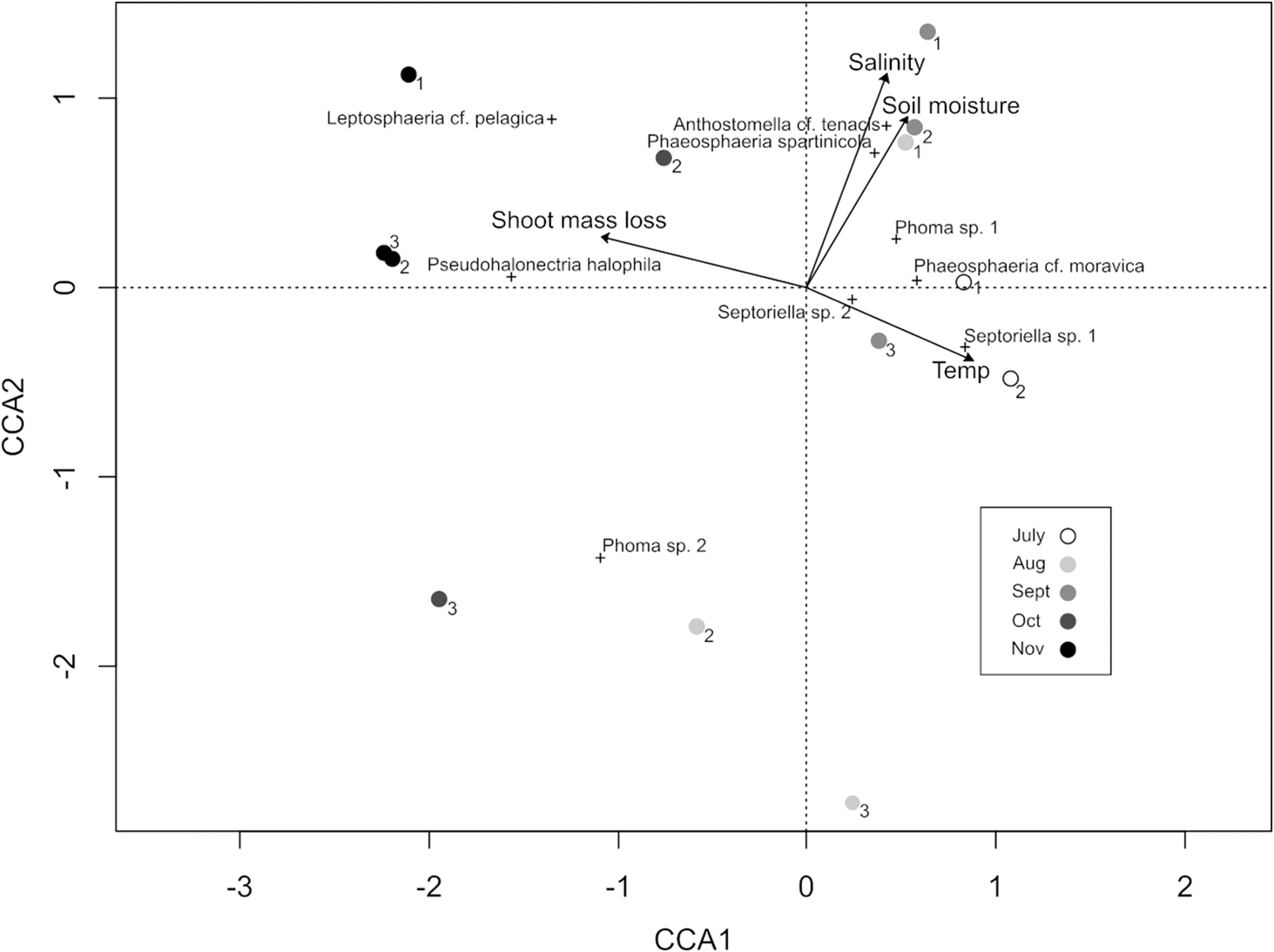
Seventeen fungi were observed fruiting in S. pumilus shoots during this study (Table 2); eight were teleomorphic and nine were anamorphic. A relatively large number of apparently undescribed species were encountered, in most cases only allowing for the identification of morphospecies within genera. The exact number of true species may be smaller than this depending on anamorph–teleomorph relationships, which were not determined in this study. Monthly richness peaked in July, when 13 species were encountered across the three sites (Supplementary Table S1). Five species were encountered in June, August, September, and October, and 2 species were encountered in November. The most abundant taxa were Septoriella sp. 1, Septoriella sp. 2, Phoma sp. 1, and Pseudohalonectria halophila. Septoriella sp. 1 was encountered only in June and July in very fresh leaves, and was present in later months only as empty pycnidia as leaf tissue became increasingly fragmented. Septoriella sp. 2 was present from June to October, but by September most pycnidia were invaded by bacteria which partially decomposed the remaining conidia. Phoma sp. 1 was encountered from June to October (but was not observed in August), and appeared fresh regardless of the state of decomposition of the leaf litter. Pseudohalonectria halophila was very commonly encountered in October and November; however, it was only in an immature state. This species seems to represent an important member of the late successional community, with the only other co-occurring fungal taxon encountered during these later months being Leptosphaeria cf. pelagica. Ten species were cumulatively found at each of the three sites, each supporting a somewhat different assemblage. Fungal species richness decreased predictably over time (Fig. 3a), displaying a significant positive correlation with shoot litterbag weight and surface temperature; however, shoot litterbag weight appears to be more important than surface temperature for structuring fungal communities (Table 1). Fungal richness and mite richness were inversely correlated (Fig. 3b). Canonical correspondence analysis showed surface temperature and shoot mass loss were the best predictors of fungal community composition (Fig. 4).
Table 2.
| Class | Family | Species |
|---|---|---|
| Dothideomycetes | Didymellaceae | Epicoccum nigrum |
| Leptosphaeriaceae | Leptosphaeria cf. pelagica | |
| Phaeosphaeriaceae | Phaeosphaeria sp. | |
| Phaeosphaeria spartinicola | ||
| Phaeosphaeria cf. moravica | ||
| Septoriella sp. 1 | ||
| Septoriella sp. 2 | ||
| Pleosporaceae | Alternaria maritima | |
| Sordariomycetes | Nectriaceae | cf. Fusarium |
| Phomatosporaceae | Phomatospora bellaminuta | |
| Pseudohalonectriaceae | Pseudohalonectria halophila | |
| Xylariaceae | Anthostomella cf. tenacis | |
| Incertae sedis | Incertae sedis | cf. Microsphaeropsis |
| cf. Mycosphaerella | ||
| Phoma sp. 1 | ||
| Phoma sp. 2 |
Fig. 3.
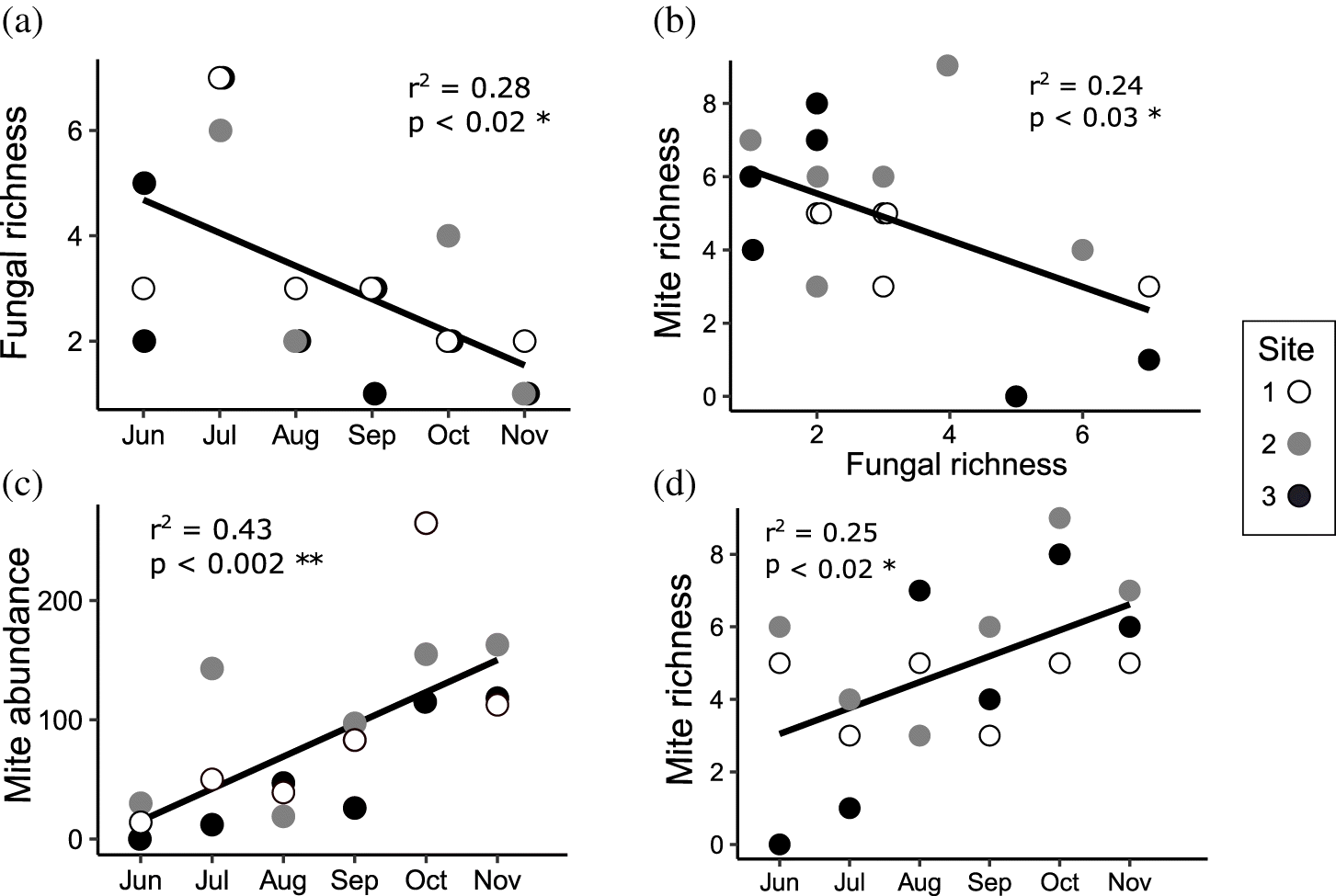
Fig. 4.
Mite community composition
Twenty-six mite species were encountered in this study (Table 3). The most species-rich group were the Trombidiform mites, represented by 15 species, followed by the Oribatid mites with 8 species. Mesostigmata, Astigmata, and Endeostigmata were each represented by 1 species. The families Erythraeidae, Tarsonemidae, and Tydeidae were the most species-rich families present. Erythraeidae was represented by two undescribed species of Abrolophus, and a species that could not be resolved to the genus level. Of the Tarsonemidae, there were two undescribed species of Steneotarsonemus, and a third species not identified to genus. Of the Tydaeidae were two undescribed species of Brachytydeus resembling B. manitobensis, and Tydeus sp. similar to the European species T. kochi. The most abundant Oribatid mites were Pelopsis bifurcatus, Diapterobates notatus, and Hermannia subglabra, all of which have previously been reported from Nova Scotia salt marshes (Behan-Pelletier et al. 1987).
Table 3.
| Order | Family | Species |
|---|---|---|
| Astigmata | Acaridae | Acotyledon cf. sokolovi |
| Mesostigmata | Blattisociidae | Cheirsoseius sp. |
| Oribatida | Ameronothridae | Ameronothrus lineatus |
| Brachychthoniidae | Brachychthoniidae sp. | |
| Cepheidae | Ommatocepheus sp. | |
| Hermanniidae | Hermannia subglabra | |
| Humerobatidae | Diapterobates notatus | |
| Hydrozetidae | cf. Hydrozetes | |
| Phthiracaridae | Phthiracarus sp. | |
| Punctoribatidae | Pelopsis bifurcatus | |
| Trombidiformes | Eupodidae | Pseudoeupodes sp. |
| Erythraeidae | Abrolophus sp. 1 | |
| Abrolophus sp. 2 | ||
| Erythraeidae sp. | ||
| Halacaridae | Isobactrus levis | |
| Iolinidae | Microtydeus sp. | |
| Microdispidae | Microdispidae sp. | |
| Nanorchestidae | Nanorchestes sp. | |
| Triophtydeidae | Triophtydeus immanis | |
| Stigmaeidae | Cheylostigmaeus sp. | |
| Tarsonemidae | Steneotarsonemus cf. badulini | |
| Steneotarsonemus sp. 2 | ||
| Tarsonemidae sp. | ||
| Tydeidae | Brachytydeus sp. 1 | |
| Brachytydeus sp. 2 | ||
| Tydeus cf. kochi |
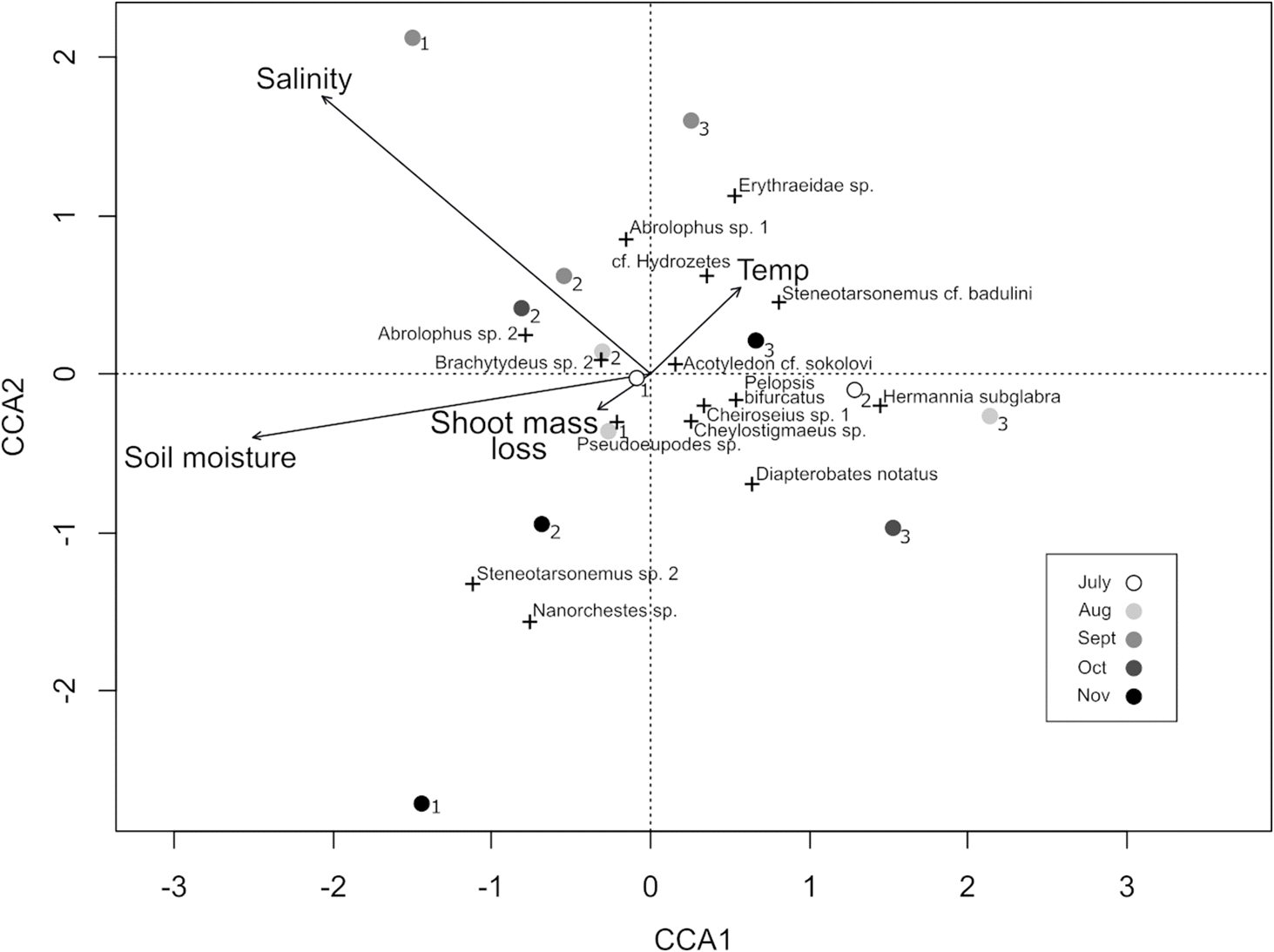
Across all three sites, 1534 mites were collected, 977 of which were immature instars. In the early months, abundance was very low but rose steadily throughout the study (Fig. 3c), showing a strong temporal trend but in the opposite direction as predicted. Richness followed a similar trend to abundance (Fig. 3d). Shannon diversity increased with time, but this was not significant (p = 0.1, R2 = 0.1) because of low absolute abundance in June, resulting in higher evenness and consequently higher Shannon diversity values. Omitting the month of June results in a significant correlation (p < 0.03, r2 = 0.3). Notably, Isobactris levis was the only member of the obligate marine family Halacaridae encountered, observed only in June. Pseudoeupodes sp. and Nanorchestes cf. amphibius were fairly abundant in October and November, but not observed in earlier months. In total, 21 species were observed at site 2, 15 species at site 1, and 14 species at site 3. Shoot litterbag weight was the best predictor of mite diversity (Table 1), while canonical correspondence analysis revealed shoot mass loss and surface temperature were the best predictor of mite community composition, with late-season species clustering together and most of the remaining species clustering near the origin (Fig. 5).
Fig. 5.
Discussion
This study assessed the biodiversity of two decomposer communities associated with the decomposition of Sporobolus pumilus leaf litter in a Nova Scotia salt marsh. A litterbag study was employed to quantify decomposition rate of shoots and roots, and mite and fungal communities were surveyed from litterbag contents. Decomposition of aboveground material occurred more rapidly than belowground material, with aboveground bags losing 44% of initial mass by November and belowground bags losing 32%. Aboveground decomposition occurred more rapidly than in other studies, such as Foote and Reynolds (1997) and Windham (2001), both of which found approximately 30% mass loss after 6 months using litterbags of similar design to this study. Root decomposition of S. pumilus is less frequently studied than shoots and leaves, and root decomposition rates appear to be highly variable in response to variation in sediment type and nitrogen content (Conn and Day 1997). Seventeen fungal species were encountered, with species richness peaking in July and falling rapidly by November. By contrast, 26 mite species were encountered, occurring at relatively low abundance and richness in the summer months and peaking in October. Fungal richness and mite diversity across sites was most strongly associated with shoot litterbag weight and surface temperature, while salinity and moisture were not associated.
Site characteristics and decomposition
Aboveground litter bags were found to decompose more rapidly than belowground litterbags, supporting our first hypothesis. The dense, waterlogged sediments may explain why roots decomposed more slowly than aboveground material. Clay-rich salt marsh sediments are generally hypoxic, especially with periodic tidal inundation (Colmer et al. 2013). Structural differences also exist between root and shoot material which might explain differences in their decomposition rates. Sporobolus roots are also relatively high in lignocellulose, which is resistant to decomposition (Hodson et al. 1984); however, Ferrer et al. (2020) found that marine fungi are surprisingly efficient at degrading this substrate. The compositional differences between litter types and the implications for decomposition rates have also been observed between different species of salt marsh grasses. Frasco and Good (1982) found that S. pumilus decomposes more slowly than the relatively nitrogen-rich S. alterniflorus, but they also suggested the presence of aerenchyma in the stems of S. alterniflorus increase its surface area, potentially facilitating decomposition.
Differences in decomposition among the three salt marsh sites were less pronounced. We found that site 3 exhibited higher decomposition rates as well as a lower soil moisture level, although there was no significant correlation between decomposition rates and soil moisture. This may be because this site is geographically higher than sites 1 and 2, resulting in less frequent tidal inundation. Similar results were observed for the tea bag decomposition assay, where decomposition rate (k) was highest at site 3. Overall, tea bag mass loss was similar to that reported by Marley et al. (2019) at two Scottish salt marshes. Soil moisture is positively correlated with decomposition in both the laboratory (Lee et al. 2014) and the field (Cortez 1998; A’Bear et al. 2014), sometimes exhibiting a nonlinear relationship where decomposition rate decreases at very high-water contents (Roper 1985; Wang et al. 2019), although this may only occur if soils become anaerobic (Day 1983). In a study of decomposition rates of S. pumilus and S. alterniflorus, Halupa and Howes (1995) found moisture was the most important factor regulating decomposition rates of aboveground material, which increased exponentially with increasing moisture.
The only environmental variable measured in this study that significantly correlated with aboveground decomposition rates was surface temperature. Temperature and decomposition rates were both highest in the summer months. Temperature positively impacts decomposition rates either independently or in combination with moisture levels (Kirschbaum 1995; Ise and Moorcroft 2006). Soil salinity was not correlated with aboveground or belowground decomposition rates. Salinity at sites 1 and 2 were on average above 35 ppt, the average salinity level of sea water. This is likely due to infrequent tidal inundation of the high marsh, followed by periods of evaporation. Rain can have a desalinating effect on the soils, where high marsh habitats can exhibit salinities only somewhat higher than terrestrial habitats or salinities at higher concentration than sea water depending on inundation frequency and rainfall frequency (Callaway et al. 1990; Xin et al. 2017). Fungal activity is inhibited by high salt concentration (Malik et al. 1979; Rath et al. 2019), but the impact of soil salinity on decomposition rates in salt marshes is variable (Hemminga et al. 1991; Qu et al. 2019).
Fungal richness
Shoot decomposition rate was the best predictor for both fungal and mite richness; however, only fungal richness was positively correlated with shoot decomposition rate, partially supporting hypothesis 2. Temporal changes in fungal community composition were more likely tied to the decomposition state of the grass rather than to seasonality due to the substrate specificity observed in many of the species encountered. For example, Septoriella sp. 2 and Phaeosphaeria cf. moravica principally developed in shoots, whereas Septoriella sp. 1 was exclusively found in leaves, and the occurrence of these substrate-specific species coincided noticeably with the decomposition state of the litter. Leaf tissues fragmented more readily than stems as decomposition progressed, and the remaining leaf tissue from August–November mainly supported old, empty fungal fruiting bodies. In later months the only fungi present were those that fruit on stems rather than leaves, as stems were less fragmented, with the two Phoma spp. uncommonly occurring in the fragmented leaf tissue.
Fungal succession has been studied in another salt marsh grass, S. alterniflorus, where community composition was also found to be temporally variable (Gessner 1977), and species richness was highest shortly after substrate tissue death. Gessner (1977) also noted a spatial organization of species, with aerial tissues supporting “terrestrial” species and lower tissues hosting more “marine” species that often produce specialized spores bearing appendages for aquatic dispersal. A similar pattern of vertical stratification was observed in fungal communities of another salt marsh plant, Phragmites australis (Van Ryckegem et al. 2007). The only spatial partitioning of species noted in this study was between stems and leaves, and no such vertical stratification was observed. S. pumilus plants produce a much narrower stem than S. alterniflorus and P. australis, which leads to plants often becoming windswept or knocked over. This could result in a less pronounced difference in inundation frequency between upper and lower plant tissues. Also, plants that occur in the high marsh are inundated less frequently than plants of the low marsh.
Of the 17 fungal morphospecies present, only 4 were hyphomycetous. Of these, two were found only once (Trichoderma sp., Epicoccum nigrum), and one was only observed as loose spores in sediment attached to plant tissues (Fusarium sp.), suggesting that it may grow on some substrate other than S. pumilus. The production of spores or conidia in an enclosed structure (e.g., pseudothecia, pycnidia etc.) is the most representative reproductive strategy for fungi on S. pumilus, which reflects the general trend for marine fungi as a whole. Hyde et al. (2000) reported 444 marine species worldwide, of which only 56 are hyphomycetous or basidiomycetous, both of which are typically characterized by exposed sporogenesis (Kohlmeyer and Kohlmeyer 1979). Enclosed reproductive structures immersed in the substrate tissues may be the preferable reproductive strategy for fungi in S. pumilus as they would shelter the sporogenous cells from most abrasive forces caused by water currents or by friction between adjacent plants as they are blown by wind.
Fungal diversity from salt marsh inhabiting Sporobolus spp. have received very little attention in Canada. Another study that surveyed fungal diversity associated with Sporobolus spp. in the Wolfville salt marsh reported 18 species from S. alterniflorus and 3 species from S. pumilus (Boland and Grund 1979), none of which from the latter species were found in the present study. A recent study of arbuscular mycorrhizal fungi from S. pumilus reported a single species commonly found colonizing root material (d’Entremont et al. 2018). Additionally, Miller and Whitney (1983) surveyed fungi from living S. alterniflorus material in Atlantic Canada, reporting 26 predominantly terrestrial generalist species.
Mite richness
Contrary to expectations, mite richness was negatively correlated with fungal richness, refuting our third hypothesis. This was due to mite richness increasing gradually over the course of the season, rather than following fungal community dynamics as predicted. As fungi are directly responsible for the decomposition of the leaf litter, high fungal richness in the early months is understandable. Mite abundance and richness were lowest during June and July when fungal richness was at its highest, suggesting spores and conidia of S. pumilus associated fungi may not be an important food source for salt marsh mites despite the abundance of S. pumilus litter in the high marsh. The increasing mite abundance over time may be explained by the following.
1.
Leaf litter becomes palatable to mites only after it has been sufficiently decomposed by fungi and bacteria. Some larger Oribatid mites have been observed to use higher plants as a major food source (Kaneko 1988), belonging to a feeding guild known as macrophytophages, most widely represented by the Ptychoidea (Schuster 1956). Macrophytophagous taxa were mostly absent from the study although Hermannia subglabra, a relatively abundant Oribatid mite in later months, is possibly a macrophytophage given that a congeneric has been reported as feeding primarily on woody tissue in juvenile stages (Wallwork 1958). A drawback of this explanation is that much of the increased mite abundance over time is driven by small-bodied taxa that probably lack the mouthparts capable of feeding directly on leaf litter such as Nanorchestes sp. and Brachytydeus sp. 2. The latter species was the most abundant mite encountered in this study, and gut contents were sometimes observed in cleared specimens. Gut contents were found to contain large quantities of intact fungal spores. Analyzing gut contents was not a focus of this study and so only a few individuals were examined in this respect, however it may be that this species is too small to possess chelicerae powerful enough to macerate plant cell walls or even fungal cell membranes.
2.
Mites mostly do not rely on S. pumilus litter and its associated fungi for food. Other food sources may be more important such as algae. Leaf litter in this study became increasingly encrusted with cyanobacteria, and diatoms are abundant in surface sediments (Laird and Edgar 1992). Fungi from sources other than S. pumilus may also be important food items. A slide-mounted specimen of Brachytydeus sp. 2 had gut contents containing approximately 100 ascospores most likely belonging to an unidentified member of the Thelebolaceae, a family of coprophilous ascomycetes. Also, one slide-mounted specimen of Cheiroseius sp., a predatory mite, was found with Phaeotrichum sp. spores attached to one of its hind tarsi. This fungal genus occurs exclusively in rodent dung. Additionally, cleistothecia of this fungus were occasionally encountered in the ethanol-containing Tullgren-Berlese mite extractions during this study. This suggests dung may be frequented by some salt marsh mites as a food source. Three species of Phaeotrichum are reported from eastern Canada: P. hystricinum and P. cylindrosporum from porcupine dung and P. circinatum from lemming dung. The Phaeotrichum species encountered is possibly an undescribed species from mouse or vole dung, as neither porcupines nor lemmings are likely to be found in the Wolfville salt marsh. The impacts of small rodents and their dung on adjacent communities does not appear to have been studied.
3.
Mite diversity may have been responding to the accumulation of fresh leaf litter as the season progressed. Richness and abundance peaked in November, which is approximately the time of year when the shoots and leaves of S. pumilus die off before winter. The litterbag experiment employed in this study used overwintered leaf litter from the previous growing season, so the apparent disconnect between litter decomposition and mite diversity may have instead been due to the mites exhibiting a preference for fresh leaf litter and the fungi associated with it. Given that the litterbags are permeable to mites, changes to litter quantity and quality outside the litterbags cannot be ruled out as an important factor increasing mite richness and abundance overall. Mites obtained from litterbags possibly represent a sample of the entire salt marsh litter-inhabiting mite community rather than mites that have a preference for overwintered leaf litter. Additionally, there may be other factors related to seasonality that mites may be utilizing.
Conclusions
This study quantified aboveground and belowground decomposition rates for S. pumilus, the dominant salt marsh grass in Nova Scotia salt marshes. Shoots and leaves on average lost 44% of their mass after 6 months, and roots lost 32%. This study also provides a first examination of the temporal dynamics of two important decomposer groups associated with S. pumilus in Canada. Fungal and mite communities both exhibited pronounced compositional changes over the course of the experiment, with the two communities appearing most active at opposite ends of the decomposition process. Future studies into relationships between salt marsh fungi and mites should consider the role of dung as a source of novel species and as an important source of nutrients entering salt marsh sediments.
Acknowledgments
BM received generous support from the Arthur Irving Family Foundation Graduate Scholarship, Acadia University. AKW and BM gratefully acknowledge research support from the Kenneth A. Harrison Memorial Mycological Fund, Acadia University; AKW and GK acknowledge NSERC Discovery Grants (No. NSERC-2017-04325 and No. NSERC-2015-04323, respectively). We thank J.C. López-Gutiérrez, R. Browne, A. Belliveau and Acadia’s E.C. Smith Herbarium, D. Divanli, C. Vail, C. Cooper, S. Adams, A. Bunbury-Blanchette, and K. Sampson for lab and field assistance.
References
A’Bear AD, Jones TH, Kandeler E, and Boddy L. 2014. Interactive effects of temperature and soil moisture on fungal-mediated wood decomposition and extracellular enzyme activity. Soil Biology and Biochemistry, 70: 151–158.
Ainsworth GC, Sparrow FK, and Sussman AS. 1973. The fungi: an advanced treatise: vol. IVA. Academic Press, New York, New York, 621 p.
Alongi D. 2020. Carbon balance in salt Marsh and Mangrove ecosystems: a global synthesis. Journal of Marine Science and Engineering, 8(10): 767.
Anderson DR, Burnham KP, and White GC. 1998 Comparison of Akaike information criterion and consistent Akaike information criterion for model selection and statistical inference from capture-recapture studies. Journal of Applied Statistics, 25(2): 263–282.
Balogh J. 1972. The Oribatid genera of the world. Akadémiai Kiadó, Budapest, Hungary. 220 p.
Barr ME. 1990. Prodromus to nonlichenized, pyrenomycetous members of class Hymenoascomycetes. Mycotaxon, 39: 43–184.
Barr ME. 2004. Prodromus to class Loculoascomycetes. Hamilton I. Newell Inc, Amherst, MA. 168 p.
Behan-Pelletier VM, and Lindo Z. 2019. Checklist of oribatid mites (Acari: Oribatida) of Canada and Alaska. Zootaxa, 4666(1): 1–180.
Behan-Pelletier VM, Lindquist EE, and Smith IM. 1987. Subclass Acari (mites and ticks). In The insects, spiders and mites of Cape Breton highlands National Park. Edited by Lafontaine, JD, Allyson, S, Behan-Pelletier, VM, Borkent, A, Campbell, JM, Hamilton, KGA, et al. Agriculture Canada Biosystematics Research Centre, Ottawa, Canada, 301p.
Benner R, Newell SY, Maccubbin AE, and Hodson RE. 1984. Relative contributions of bacteria and fungi to rates of degradation of Lignocellulosic Detritus in salt-Marsh sediments. Applied and Environmental Microbiology, 48(1): 36–40.
Berg MP, and Bengtsson J. 2007. Temporal and spatial variability in soil food web structure. Oikos, 116(11): 1789–1804.
Boesch DF, and Turner RE. 1984. Dependence of fishery species on salt marshes: the role of food and refuge. Estuaries, 7(4): 460–468.
Boland GJ, and Grund DW. 1979. Fungi from the salt marshes of Minas Basin, Nova Scotia. Proceedings of the Nova Scotia Institute of Science, 29: 393–404.
Callaway RM, Jones S, Ferren WR Jr, and Parikh A. 1990. Ecology of a mediterranean-climate estuarine wetland at Carpinteria, California: plant distributions and soil salinity in the upper marsh. Canadian Journal of Botany, 68(5): 1139–1146.
Chapman V. 1937. A note on the salt marshes of Nova Scotia. Rhodora, 39(459): 53–57.
Chmura GL, Anisfeld SC, Cahoon DR, and Lynch JC. 2003. Global carbon sequestration in tidal, saline wetland soils. Global Biogeochemical Cycles, 17(4).
Colmer TD, Pedersen O, Wetson AM, and Flowers TJ. 2013. Oxygen dynamics in a salt-marsh soil and in Suaeda maritima during tidal submergence. Environmental and Experimental Botany, 92: 73–82.
Conn CE, and Day FP Jr. 1997. Root decomposition across a barrier island chronosequence: litter quality and environmental controls. Plant and Soil, 195: 351–364.
Cortez J. 1998. Field decomposition of leaf litters: relationships between decomposition rates and soil moisture, soil temperature and earthworm activity. Soil Biology and Biochemistry, 30(6): 783–793.
Danger M, Cornut J, Elger A, and Chauvet E. 2012. Effects of burial on leaf litter quality, microbial conditioning and palatability to three shredder taxa. Freshwater Biology, 57(5): 1017–1030.
Darnell TM, and Smith EH. 2004. Avian use of natural and created salt Marsh in Texas, USA. Waterbirds, 27(3): 355–361.
Day FP. 1983. Effects of flooding on leaf litter decomposition in microcosms. Oecologia, 56(2): 180–184.
Deegan LA, Hughes JE, and Rountree RA. 2000. Salt Marsh ecosystem support of Marine transient species. In Concepts and controversies in Tidal Marsh ecology. Edited by Weinstein, MP, and Kreeger, DA Springer, Dordrecht, Netherlands. p. 875.
d’Entremont TW, López-Gutiérrez JC, and Walker AK. 2018. Examining Arbuscular Mycorrhizal Fungi in Saltmarsh Hay (Spartina patens) and Smooth Cordgrass (Spartina alterniflora) in the Minas Basin, Nova Scotia. Northeastern Naturalist, 25(1): 72–86
Dindal DL. 1990. Soil biology guide, Wiley Interscience, Canada. 1349 p.
Ferrer A, Heath KD, Canam T, Flores HD, and Dalling JW. 2020. Contribution of fungal and invertebrate communities to wood decay in tropical terrestrial and aquatic habitats. Ecology, 101(9): e03097.
Foote AL, and Reynolds KA. 1997. Decomposition of saltmeadow cordgrass (Spartina patens) in Louisiana coastal marshes. Estuaries, 20: 579–588.
Frasco BA, and Good RE. 1982. Decomposition dynamics of Spartina alterniflora and Spartina patens in a New Jersey Salt Marsh. American Journal of Botany, 69(3): 402–406.
Gessner RV. 1977. Seasonal occurrence and distribution of Fungi associated with Spartina alterniflora from a Rhode Island Estuary. Mycologia, 69(3): 477–491.
Gilyarov МС. 1975. A key to the soil-inhabiting mites (Sarcoptiformes). Nauka, Moscow, Russia. 492 p.
Gilyarov МС. 1977. A key to the soil-inhabiting mites (Mesostigmata). Nauka, Moscow, Russia. 720 p.
Gilyarov МС. 1978. A key to the soil-inhabiting mites (Trombidiformes). Nauka, Moscow, Russia. 272 p.
Guerra CA, Heintz-Buschart A, Sikorski J, Chatzinotas A, Guerrero-Ramírez N, Cesarz S, et al. 2020. Blind spots in global soil biodiversity and ecosystem function research. Nature Communications, 11(1): 3870.
Halupa PJ, and Howes BL. 1995. Effects of tidally mediated litter moisture content on decomposition of Spartina alterniflora and S. patens. Marine Biology, 123(2): 379–391.
Hättenschwiler S, Tiunov AV, and Scheu S. 2005. Biodiversity and litter decomposition in terrestrial ecosystems. Annual Review of Ecology, Evolution, and Systematics, 36: 191–218.
Hemminga MA, de Leeuw J, de Munek W, and Koutstaal BP. 1991. Decomposition in estuarine salt marshes: the effect of soil salinity and soil water content. Vegetatio, 94(1): 25–33.
Hodson RE, Christian RR, and Maccubbin AE. 1984. Lignocellulose and lignin in the salt marsh grass Spartina alterniflora: initial concentrations and short-term, post-depositional changes in detrital matter. Marine Biology, 81: 1–7.
Howes BL, and Teal JM. 1994. Oxygen loss from Spartina alterniflora and its relationship to salt marsh oxygen balance. Oecologia, 97: 431–438.
Hughes RG. 2004. Climate change and loss of saltmarshes: consequences for birds. Ibis, 146(s1): 21–28.
Hyde KD, Sarma VV, and Jones EBG. 2000. Morphology and taxonomy of higher marine fungi. In Marine mycology: a practical approach. Edited by Hyde, KD, and Pointing, SB. Fungal Diversity Press, Hong Kong. p. 377.
Ise T, and Moorcroft PR. 2006. The global-scale temperature and moisture dependencies of soil organic carbon decomposition: an analysis using a mechanistic decomposition model. Biogeochemistry, 80(3): 217–231.
Jones EBG, Sakayaroj J, Suetrong S, Somrithipol S, and Pang KL. 2009. Classification of marine Ascomycota, anamorphic taxa and Basidiomycota. Fungal Diversity, 73(1): 1–72.
Kaneko N. 1988. Feeding habits and cheliceral size of oribatid mites in cool temperate forest soils in Japan. Revue d’Écologie et de Biologie du Sol [Review of Soil Ecology and Biology], 25: 353–363.
Keuskamp JA, Dingemans BJJ, Lehtinen T, Sarneel JM, and Hefting MM. 2013. Tea Bag index: a novel approach to collect uniform decomposition data across ecosystems. Methods in Ecology and Evolution, 4(11): 1070–1075.
Kirschbaum MUF. 1995. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage. Soil Biology and Biochemistry, 27(6): 753–760.
Klironomos JN, and Kendrick WB. 1996. Palatability of microfungi to soil arthropods in relation to the functioning of arbuscular mycorrhizae. Biology and Fertility of Soils, 21(1): 43–52.
Kohlmeyer J, and Kohlmeyer E. 1979. Marine mycology. Academic Press, New York, New York. 690 p.
Krantz GW, and Walter DE. 2009. A manual of acarology. Texas Tech University Press, Lubbock, Texas. 807 p.
Krest JM, Moore WS, Gardner LR, and Morris JT. 2000. Marsh nutrient export supplied by groundwater discharge: evidence from radium measurements. Global Biogeochemical Cycles, 14(1): 167–176.
Laird K, and Edgar RK. 1992. Spatial distribution of diatoms in the surficial sediments of a New England Salt Marsh. Diatom Research, 7(2): 267–279.
Lee H, Fitzgerald J, Hewins DB, McCulley RL, Archer SR, Rahn T, et al. 2014. Soil moisture and soil-litter mixing effects on surface litter decomposition: a controlled environment assessment. Soil Biology and Biochemistry, 72: 123–132.
López-Mondéjar R, Brabcová V, Štursová M, Davidová A, Jansa J, Cajthaml T, et al. 2018. Decomposer food web in a deciduous forest shows high share of generalist microorganisms and importance of microbial biomass recycling. The ISME Journal, 12(7): 1768–1778.
Lussenhop J. 1992. Mechanisms of microarthropod-microbial interactions in soil. In Advances in ecological research vol. 23. Edited by Begon, M and Fitter, AH. Academic Press, New York, New York. p. 355.
Lyons JI, Newell SY, Brown RP, and Moran MA. 2005. Screening for bacterial–fungal associations in a south-eastern US salt marsh using pre-established fungal monocultures. FEMS Microbiology Ecology, 54(2): 179–187.
Mąkol J, and Wohltmann A. 2012. An annotated checklist of terrestrial Parasitengona (Actinotrichida: Prostigmata) of the world, excluding Trombiculidae and Walchiidae. Annales Zoologici, 62(3): 359–562.
Malik KA, Bhatti NA, and Kauser F. 1979. Effect of soil salinity on decomposition and humification of organic matter by some cellulolytic fungi. Mycologia, 71(4): 811–820.
Marley ARG, Smeaton C, and Austin WEN. 2019. An assessment of the Tea Bag index method as a proxy for organic matter decomposition in intertidal environments. Journal of Geophysical Research: Biogeosciences, 124(10): 2991–3004.
Mazerolle M. 2020. AICcmodavg: model selection and multimodel inference based on (Q)AIC(c). R package version 2.3-1. [online]: Available from cran.r-project.org/package=AICcmodavg.
Mazzoleni S, Bonanomi G, Giannino F, Rietkerk M, Dekker S, and Zucconi F. 2007. Is plant biodiversity driven by decomposition processes? An emerging new theory on plant diversity. Community Ecology, 8(1): 103–109.
Miller JD, and Whitney NJ. 1983. Fungi of the Bay of Fundy V: fungi from living species of Spartina Schreber. Proceedings of the Nova Scotian Academy of Science, 33: 75–83.
Möller I, Kudella M, Rupprecht F, Spencer T, Paul M, van Wesenbeeck BK, et al. 2014. Wave attenuation over coastal salt marshes under storm surge conditions. Nature Geoscience, 7(10): 727–731.
Newell SY, Fallon RD, and Miller JD. 1989. Decomposition and microbial dynamics for standing, naturally positioned leaves of the salt-marsh grass Spartina alterniflora. Marine Biology, 101(4): 471–481.
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, et al. 2020. Vegan: community ecology package. R package version 2.5-7. [online]: Available from CRAN.R-project.org/package=vegan.
Pollierer MM, and Scheu S. 2021. Stable isotopes of amino acids indicate that soil decomposer microarthropods predominantly feed on saprotrophic fungi. Ecosphere, 12(3): e03425.
Prescott CE, Zabek LM, Staley CL, and Kabzems R. 2000. Decomposition of broadleaf and needle litter in forests of British Columbia: Influences of litter type, forest type, and litter mixtures. Canadian Journal of Forest Research, 30(11): 1742–1750.
Qu W, Li J, Han G, Wu H, Song W, and Zhang X. 2019. Effect of salinity on the decomposition of soil organic carbon in a tidal wetland. Journal of Soils and Sediments, 19(2): 609–617.
R Core Team. 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing. [online]: Available from R-project.org/.
Rath KM, Murphy DN, and Rousk J. 2019. The microbial community size, structure, and process rates along natural gradients of soil salinity. Soil Biology and Biochemistry, 138: 107607.
Roper MM. 1985. Straw decomposition and nitrogenase activity (C2H2 reduction): Effects of soil moisture and temperature. Soil Biology and Biochemistry, 17(1): 65–71.
Schuster R. 1956. Der Anteil der Oribatiden an den Zersetzungsvorgängen im Boden. Zeitschrift für Morphologie und Ökologie der Tiere [The proportion of oribatid mites in the decomposition processes in the soil. Journal of Morphology and Ecology of Animals], 45(1): 1–33.
Seifert KA, Morgan-Jones G, Gams W, and Kendrick B. 2011. The genera of hyphomycetes. CBS-KNAW Fungal Biodiversity Centre, Utrecht, Netherlands. 997 p.
Shepard CC, Crain CM, and Beck MW. 2011. The protective role of coastal marshes: A systematic review and meta-analysis. PLoS ONE, 6(11): e27374.
Shoemaker RA, and Babcock CE. 1989. Phaeosphaeria. Canadian Journal of Botany, 67: 1500–1599.
Silva GLD, Metzelthin MH, Silva OSD, and Ferla NJ. 2016. Catalogue of the mite family Tydeidae (Acari: Prostigmata) with the world key to the species. Zootaxa, 4135(1): 1–68.
Suberkropp K, and Arsuffi TL. 1984. Degradation, growth, and changes in palatability of leaves colonized by six aquatic hyphomycete species. Mycologia, 76(3): 398–407.
Subías LS, Shtanchaeva UY, and Arillo A. 2012. Listado de los ácaros oribátidos (Acariformes, Oribatida) de las diferentes regiones biogeográficas del mundo. Monografia Electronica de Sociedad Entomologica. [online]: Available from sea-entomologia.org/gia/libros_subias.html.
Sutton BC. 1980. The coelomycetes. Fungi imperfecti with pycnidia, acervuli and stromata. Commonwealth Mycological Institute, Surrey, England. 696 p.
ter Braak CJF. 1986. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology, 67(5): 1167–1179.
Van Ryckegem G, Gessner MO, and Verbeken A. 2007. Fungi on leaf blades of phragmites Australis in a Brackish Tidal marsh: diversity, succession, and leaf decomposition. Microbial Ecology, 53(4): 600–611.
Wallwork JA. 1958. Notes on the feeding behaviour of some forest soil Acarina. Oikos, 9(2): 260–271.
Walter D, Latonas S, Byers K, and Lumley L. 2014. Almanac of Alberta Oribatida Part I, Version 2.4. [online]: Available from researchgate.net/publication/352842283_Almanac_of_Alberta_Oribatida_Part_I_Version_24.
Wang F, Kroeger KD, Gonneea ME, Pohlman JW, and Tang J. 2019. Water salinity and inundation control soil carbon decomposition during salt marsh restoration: an incubation experiment. Ecology and Evolution, 9(4):1911–1921.
Wickham H. 2016. ggplot2: elegant graphics for data analysis. [online]: Available from ggplot2.tidyverse.org.
Windham L. 2001. Comparison of biomass production and decomposition between Phragmites australis (common reed) and Spartina patens (salt hay grass) in brackish tidal marshes of New Jersey, USA. Wetlands, 21: 179–188.
Xin P, Zhou T, Lu C, Shen C, Zhang C, D’Alpaos A, and Li L. 2017. Combined effects of tides, evaporation and rainfall on the soil conditions in an intertidal creek-marsh system. Advances in Water Resources, 103: 1–15.
Yan J, Wang L, Hu Y, Tsang YF, Zhang Y, Wu J, et al. 2018. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability. Geoderma, 319: 194–203.
Young MR, Proctor HC, deWaard JR, and Hebert PDN. 2019. DNA barcodes expose unexpected diversity in Canadian mites. Molecular Ecology, 28(24): 5347–5359.
Supplementary material
Supplementary Material 1
- Download
- 14.44 KB
Information & Authors
Information
Published In

FACETS
Volume 7 • Number 1 • January 2022
Pages: 654 - 673
Editor: Alfredo Justo
History
Received: 12 November 2021
Accepted: 16 February 2022
Version of record online: 5 May 2022
Notes
This paper is part of a collection titled “The Canadian Fungal Network 2021 Annual Meeting – Connecting fungal researchers across Canada”.
Copyright
© 2022 Malloch et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper and in the Supplementary Material.
Key Words
Sections
Subjects
Authors
Author Contributions
All conceived and designed the study.
BM performed the experiments/collected the data.
All analyzed and interpreted the data.
GK and AKW contributed resources.
All drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Bruce Malloch, Gavin Kernaghan, and Allison K. Walker. 2022. Temporal diversity patterns for fungi and mites associated with decaying Sporobolus pumilus (Spartina patens) in the Minas Basin, Nova Scotia. FACETS.
7(): 654-673. https://doi.org/10.1139/facets-2021-0175
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item