A comparison of infectious agents between hatchery-enhanced and wild out-migrating juvenile chinook salmon (Oncorhynchus tshawytscha) from Cowichan River, British Columbia
Abstract
Infectious diseases are likely contributing to large-scale declines in chinook salmon stocks in the Pacific Northwest, but the specific agents and diseases involved, and the prevalences in migratory salmon, are mostly unknown. We applied a high-throughput microfluidics platform to screen for 45 infectious agents in 556 out-migrating juvenile chinook salmon, collected from freshwater (FW) and saltwater (SW) locations in the Cowichan River system on Vancouver Island, Canada, during 2014. Nineteen agents (5 bacterial, 2 viral, and 12 parasitic) were detected, with prevalences ranging from 0.2% to 57.6%. Co-infections between Candidatus Branchiomonas cysticola Toenshoff, Kvellestad, Mitchell, Steinum, Falk, Colquhoun & Horn, 2012, Paranucleospora theridion Nylund, Nylund, Watanabe, Arnesen & Kalrsbakk, 2010, and gill chlamydia, all associated with gill disease, were observed in SW samples. We detected agents known to cause large-scale mortalities in Pacific salmon (Ceratonova shasta (Noble, 1950), Parvicapsula minibicornis Kent, Whitaker & Dawe, 1977), and agents only recently reported in Pacific salmon in BC (Ca. B. cysticola, P. theridion, Facilispora margolisi Jones, Prosperi-Porta & Kim, 2012 and Parvicapsula pseudobranchicola Karlsbakk, Saether, Hostlund, Fjellsoy & Nylund, 2002). Wild and hatchery fish were most divergent in agent profiles in FW, with higher agent diversity in wild fish. Differences in prevalence largely dissipated once they converged in the marine environment, although hatchery fish may be infected by a greater diversity of agents sooner after ocean entry by virtue of their more rapid migration from nearshore to offshore environments.
Introduction
Pacific salmon consist of several iconic fish species belonging to the Oncorhynchus genus and have strong cultural, social, economic, recreational, and symbolic significance to the residents and First Nations in the Pacific Northwest (National Research Council 1996; Lichatowich and Lichatowich 2001). Pacific salmon are also considered a keystone species for maintaining both terrestrial and aquatic ecosystems through their function as live prey and the enrichment of stream productivity that occurs as their abundant carcasses decompose (Cederholm et al. 1999; Gende et al. 2002). In the past few decades, productivity of many stocks of Pacific salmon species (principally coho (Oncorhynchus kisutch (Walbaum, 1792)), chinook (Oncorhynchus tshawytscha (Walbaum, 1792)), and sockeye (O. nerka (Walbaum, 1792))) have been in decline, with marine mortality exceeding 90% (Beamish et al. 1995; Gresh et al. 2000; Beamish et al. 2012a; Peterman and Dorner 2012); early marine survival is most critical for overall marine survival (Pearcy 1992) and establishing year-class strength (Beamish and Mahnken 2001). Although no single factor can fully account for declines in Pacific salmon productivity, several factors have been proposed to contribute to reduced survival, including the effects of climate change, anthropogenic activities, infectious diseases, and predators. Climate change impacts the timing, abundance, and quality of prey in the early marine environment and the thermal experience in freshwater (FW) and saltwater (SW) environments. The potential cumulative or synergistic effects of these factors combined may be the most detrimental factor (Cohen 2012; Miller et al. 2014).
The Department of Fisheries and Oceans Canada (DFO) established a Salmonid Enhancement Program (SEP) in the late 1970s to increase the abundance of chinook, coho, and chum salmon through hatchery production. The major objective of the SEP was to increase the production of adult fish to enhance fishing opportunities along the coast (Hilborn and Winton 1993). The DFO-operated SEP hatchery in the Cowichan River started to produce chinook salmon in 1979 by capturing returning migrating adults. This hatchery released an average of 1.9 million (range: 0.5–3.2 million) juveniles per year from 2000 to 2009. However, in 2008, a year with poor ocean productivity, Beamish et al. (2012b) estimated that just over 1% of hatchery-reared juveniles and 8%–30% of wild juvenile from the Cowichan River survived their first marine summer.
Relatively poorer survival of hatchery fish in the marine environment in recent years has been reported in numerous studies, and is generally poorest in years of stressful ocean conditions and low availability of prey (Beamish et al. 2012b). Hatchery fish generally show reduced swimming ability (Bams 1967) and lower resistance to stress and disease (Salonius and Iwama 1993) than wild fish, which may contribute to their reduced survival at sea. Studies have shown that even a single generation of hatchery production can reduce the genetic fitness of wild fish (Araki et al. 2007; Araki et al. 2008). As a result, the use of enhancement hatcheries to produce large numbers of fish for exploitation has been debated, due to the potential for negative interactions between hatchery-reared and naturally spawned fish. Genetic introgression, overcrowding, competition, predation and predator attraction, and transfer of pathogens and disease are all factors that may carry negative consequences from hatchery to wild fish (Weber and Fausch 2003; Naish et al. 2007; Christie et al. 2012). Even though infectious diseases are, theoretically, considered to present higher risk in high-rearing-density environments like hatcheries, thus far there is no direct evidence that hatchery-released fish increase the transmission of infectious agents to wild populations (Naish et al. 2007; Stephen et al. 2011). However, there has also been no extensive study of the variation in infectious profiles between hatchery and wild fish in freshwater or marine environments.
Infectious disease is considered one of the factors potentially contributing to declines in wild salmon populations (Kent 2011), but the distribution and prevalence of infectious agents causing diseases at various life stages of salmon is mostly unknown (Miller et al. 2014). Infectious disease is a complex interaction between the characteristics of host, agent, and environment (Hedrick 1998), with the added complexity of changing environments for different life stages of anadromous fish.
Salmonid fish are host to many infectious agents including viruses, bacteria, fungi, protozoans, helminths, and arthropods. Infectious agents of salmonids are comparatively well studied; however, most of our knowledge is based on fish in culture, typically because of the difficulty associated with studies involving wild salmon populations in the large environmental zones they inhabit and the difficulty and lack of monitoring or surveillance capabilities to detect unusual mortalities (Kent 2011). Mortality events in wild salmon have been reported to be associated with the bacterium Aeromonas salmonicida (Lehmann & Neumann, 1896) Griffin, Snieszko & Friddle, 1953 in Atlantic Salmon (Emmerich 1894), C. shasta (Fujiwara et al. 2011; Hallett et al. 2012), Myxobolus cerebralis Hofer 1903 (Nehring and Thompson 2001), and Ichthyophonus hoferi Plehn & Mulsow, 1911 (Jones and Dawe 2002; Kocan et al. 2004) in Pacific salmon, and the parasites Ichthyophthirius multifiliis Fouquet, 1876 and P. minibicornis in pre-spawning BC sockeye salmon (Traxler et al. 1998; Bradford et al. 2010).
Juvenile salmon transforming from parr to smolt, and smolts moving from freshwater to marine environments, are likely to be at higher risk of contracting infections than any other life stages (Hoar et al. 1997). However, few studies have reported the monitoring of multiple infectious agents in migratory juvenile salmon, and most that do include only a handful of agents, mostly parasites (Kent et al. 1998; Arkoosh et al. 2004; Sandell 2010; Van Gaest et al. 2011). In the present study, we applied a microfluidics quantitative (q)PCR platform to ascertain the distribution, prevalence, and abundance of 45 infectious agents (bacteria, viruses, and microparasites) known or suspected to infect and cause disease in salmon. The objectives of this study were to describe the prevalence and spatiotemporal distribution, over a short time scale, during the transition from the FW to the SW phase, for each of the infectious agents in juvenile chinook salmon from the Cowichan River, and to compare differences in the prevalence and diversity of infectious agents between hatchery and wild fish. We used Cowichan chinook salmon, as the Cowichan River system has been used by researchers as a model and indicator of health and productivity for similar aquatic systems in southern British Columbia (Beamish et al. 2012b; Riddell et al. 2013), it has both hatchery and wild fish, and the juveniles from this system stay in the bay longer than many other stocks, allowing the capture of temporal variability in infectious agent distribution.
Materials and methods
For consistency, we defined all fish developed from eggs and spawned naturally in the river bed as “wild fish”. Similarly, all fish propagated artificially from eggs and milt from the group of spawners, maintained in a controlled hatchery environment until release, and identified by an adipose fin clip, were defined as “hatchery fish”. The adipose fin clip rate for Cowichan hatchery fish in 2014 was 94.5%, meaning that 5.5% of hatchery fish could be misclassified as wild.
Sampling
Hatchery and wild juvenile Cowichan chinook salmon smolts were sampled in their FW natal locations (Cowichan hatchery and Cowichan River, respectively) and SW locations (nearshore—along the shore of the bay, and bay) during their early migration to marine life in spring and summer of 2014 (See Fig. 1 for study location and Table 1 for sampling design). In 2014, smolts were released from the Cowichan hatchery on 23 April and 13 May and were collected at the hatchery on their release day. Wild fish were sampled downstream of the hatchery using a Rotary Screw Trap on 22 April and 13 May 2014, before they would have come into contact with migratory hatchery fish (which tend to move quickly downstream to the marine environment). Weight and fork length measurements of fish were recorded at the time of sampling, and each fish was assigned a unique identifier. The sampled fish were either dissected on site and tissue samples preserved as per the protocol described below, or were frozen and dissected later at the Pacific Biological Station, Nanaimo, British Columbia. The animal care and use protocol for this work was approved by the DFO Pacific Region Animal Care Committee (Animal Use Protocol Number: 13-008).
Fig. 1.
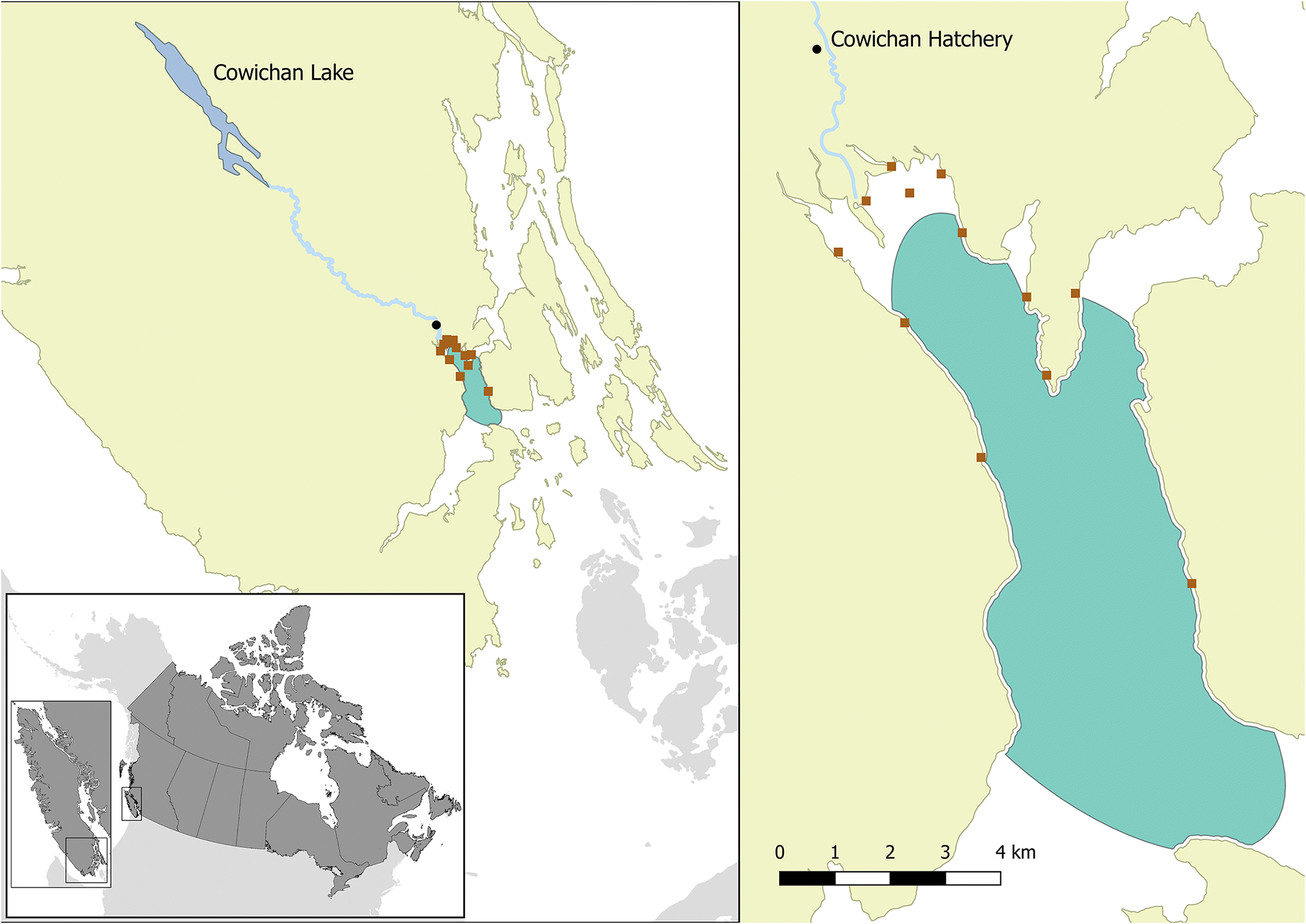
Table 1.
| Hatchery fish (No.) | Wild fish (No.) | |||||
|---|---|---|---|---|---|---|
| Sampling month | Hatchery | Nearshore | Bay | River | Nearshore | Bay |
| April | 4 | 10 | — | 27 | 14 | — |
| May | 30 | 29 | 33 | 64 | 54 | 31 |
| June | — | — | 27 | — | 33 | 93 |
| July | — | — | 18 | — | — | 80 |
| September | — | — | — | — | — | 9 |
Nearshore sampling was conducted using a beach seine from 17 April to 18 June 2014, with approximately weekly sampling intervals. Sampling locations are identified in Fig. 1. In large catches (>30 fish), a random sub-sample of 30 individuals was collected for the survey. All collected chinook salmon were given a unique identifier, weight and fork length were measured, and the presence/absence of an adipose fin was recorded to differentiate its origin as wild or hatchery.
The majority of the sampling events in the bay were conducted with a small mesh purse seine from a commercial trawler approximately every second week from 8 May to 23 July 2014. Additional sampling events in the bay in the months of June and September involved the use of a large mid-water trawl adapted to retain juvenile salmon (Beamish et al. 2000; Sweeting et al. 2003). Due to the size of the trawling vessels, these sampling events were limited to the outer reaches and associated waters outside Cowichan Bay. Live chinook salmon caught in the nets were transferred to tanks on board the trawlers for verification of the species and presence of adipose fin clips. A subsample of up to 30 chinook salmon was collected, and all fish were euthanized and either processed on the trawler or frozen and processed later at the Pacific Biological Station. The remaining live catch was released.
For our analyses, all wild chinook salmon collected within 3 km of the Cowichan River (inside Cowichan Bay) were assumed to have originated from the Cowichan River, as DNA stock identification over multiple years has shown that, in this area, virtually all fish collected in the spring and early summer are of Cowichan River origin. All chinook salmon collected outside of this 3 km region were assigned a stock of origin based on DNA analyses (Beacham et al. 2006), and fish with a >50% probability of originating from the Cowichan River, based on their DNA results, were included in our study; previous analyses of coded-wire tag data have shown a high concordance in the identification of Cowichan chinook, at a 50% probability level (C. Neville, personal observation, 2016)).
Tissue preparation and molecular assays
Fish samples were examined for 45 infectious agents (12 viruses, 11 bacterial pathogens, and 22 microparasites) using 46 qPCR assays (run in duplicate) and a single host reference gene (Table S1) run on the Fluidigm BioMark™ HD microfluidics qPCR platform (Fluidigm Corporation, San Francisco, California, USA). Dynamic arrays containing assays and samples performed qPCR reactions for 96 assays in 96 samples at once (9216 reaction wells). The qPCR assays targeted infectious agents known or hypothesized to be linked with diseases in salmon worldwide (Miller et al. 2014; Miller et al. 2016). Some of these agents are known to be endemic in BC, some were previously reported in other salmon species but not in Pacific salmon, and others have been recently described in association with emerging salmon diseases but have not yet been assessed in BC salmon. The analytical performance of individual assays analyzed on the BioMark platform has been fully evaluated and reported (Miller et al. 2016). Note that like any other qPCR technique the amplification of the genetic material of infectious agents in the samples cannot be equated to the detection of disease, only the presence and abundance of infectious agents.
Tissue preparation, RNA extraction, and qPCR assays were performed using previously described methods and protocols (Miller et al. 2011; Jeffries et al. 2014; Miller et al. 2014; Miller et al. 2016; Bass et al. 2017; Di Cicco et al. 2017). Briefly, for field dissections, small sections of tissue samples (from five tissues: brain, liver, heart, gill, and head kidney) were obtained aseptically from sampled fish and preserved in RNAlater (Qiagen, Germantown, Maryland, USA) for 24 h at 4 °C, and then frozen at −80 °C until RNA extraction. For dissections of frozen fish in the laboratory, tissues were placed directly in TRI reagent (Ambion Inc., Austin, Texas, USA) homogenization tubes.
Tissue samples were homogenized in TRI reagent to facilitate dual (total) RNA and DNA extractions from the same samples. 1-bromo-3-chloropropane was added to the homogenate and aliquots of the aqueous phase (for RNA) and organic/interphase (for DNA) were pipetted into 96-well plates. Equal volumes of aqueous phase and organic/interphase for each tissue were mixed for separate RNA and DNA extraction. RNA was extracted using the Magmax™-96 for Microarrays RNA kit (Ambion Inc., Austin, Texas, USA) with a Biomek NXP™ (Beckman-Coulter, Mississauga, Ontario, Canada) automated liquid-handling instrument, using the “spin method”, following the manufacturer’s instructions. Quantity and purity of extracted RNA were examined by measuring the A260/A280 with a Qubit 2.0 fluorometer (Invitrogen, Carlsbad, California, USA) and normalized to 62.5 ng/μL. Extracted RNA (1 μg) was reverse transcribed to cDNA (SuperScript VILO master mix kit; Invitrogen, Carlsbad, California, USA) as per the manufacturer’s instructions. DNA was extracted from the organic/interphase layer using the TNES-6U method following the Qiagen BioSprint protocol. For BioMark analysis, DNA and cDNA were mixed in equal proportions to maximize our ability to detect a wide array of infectious agents (including RNA and DNA viruses).
As the BioMark platform uses a very small assay volume (7 nL), a pre-amplification step was required. 1.3 μL of the DNA/cDNA mix from each pooled tissue sample was pre-amplified with 0.2 μmol·L−1 of 47 corresponding primer pairs (46 assays and a reference gene) in a 5-μL reaction volume using TaqMan Preamp Master Mix (Life Technologies, Carlsbad, California, USA), according to the BioMark protocol. After pre-amplification, unincorporated primers were removed using Exo-SAP-IT™ (Affymetrix, Santa Clara, California, USA), following the manufacturer’s guidelines, and then diluted 1:5 in DNA Suspension Buffer (TEKnova, Hollister, California, USA).
The 96.96 dynamic array, which contains 9216 wells for independent assessments of 96 assays against 96 samples (no multiplexing), was run as described by Miller et al. (2016). A 5 μL mixture was prepared for each sample, which contained 1 × TaqMan Universal Master Mix (No UNG), 1 × GE Sample Loading Reagent (Fluidigm PN 85000746), and the diluted pre-amplified cDNA from that sample. Separate assay mixes (5 μL) were prepared for each infectious agent, with 1 × of the TaqMan qPCR assay (infectious agent probe in FAM-MGB and artificial plasmid construct (APC) probe (see below) in NED-MGB, 10 μmol·L−1 of primers, and 3 μmol·L−1 of probes) and 1 × Assay Loading Reagent (Fluidigm PN 85000736). Negative controls included three negative processing controls for RNA/DNA extraction, two no template controls for template enrichment (described above), two cDNA (no reverse transcriptase) controls, and two no template controls for PCR. Positive controls included duplicates of a pooled sample from the cDNA/DNA for all fish used in the study, an endogenous reference gene to assess RNA quality, and five serial dilutions of APC clones for all assays to both assess assay integrity and to calculate the copy number of each detected agent; APC clones were loaded last to minimize the potential for contamination. The APC clones were synthesized and cloned sequences of the amplicon for each assay contained an “extra” probe sequence so that potential contamination of high concentration APCs in sample wells could be identified. In all, controls took up 16 sample wells, with the remaining 80 sample wells containing DNA/cDNA from individual fish. Samples were mixed and loaded into the dynamic array by the IFC HX controller (Fluidigm Corporation, San Francisco, California, USA), and the array was transferred to the BioMark instrument and processed using the GE 96×96 Standard TaqMan program for qPCR, which included an initial hot start followed by 40 cycles at 95 °C for 15 s and 60 °C for 1 min (Fluidigm Corporation, San Francisco, California, USA).
Cycle threshold (CT) values were calculated using the BioMark qPCR analysis software (Fluidigm Corporation, San Francisco, California, USA). Individual assays were evaluated for contamination, agreement between duplicates, and abnormal curve shapes. The real-time qPCR results were exported as a table view comma-separated values (csv) file and the average of the duplicated samples was calculated. Samples amplifying products from only one duplicate were treated as inconclusive and removed from analysis (for that assay).
Based on the limit of detection (LOD) for each infectious agent, as reported in the performance evaluation report for the BioMark platform (Miller et al. 2016), samples were categorized as positive only if the average of the duplicate samples was below the assay-specific LOD. The LOD is defined as the estimated CT number under which a given assay is expected to provide true positive results 95% of the time. It should be noted that the very stringent (95%) LOD used in this study (27 CT or fewer, depending on the assay) is a conservative estimate, because samples with CT <30, but not below the 27 threshold, were likely true positives. These individuals would have contained very low concentrations (1–10 copies/μL). For load analysis, we used the formula log10 (copy number + 1) for each sample and infectious agent combination.
Statistical analysis
Statistical analyses were performed using Stata (Release 14.1; StataCorp, College Station, TX, USA, 2015) and R Statistical software (R Core Team 2015). We used three approaches to analyze the data: (1) descriptive analyses to report the prevalence and load of each infectious agent and characteristics of sampled fish; (2) logistic regression models to describe the variability in the prevalence of infectious agents; and (3) quantitative models, namely multivariate and regression analyses, to describe the infectious agents’ diversity and coexistence patterns.
Descriptive analyses
Descriptive analyses were performed by summarizing infectious agent prevalence and load data. The prevalence of each of the infectious agents was defined as the number of qPCR positive fish divided by the total number of samples tested, expressed as a percentage. The spatiotemporal patterns of the prevalence of individual infectious agents were described for the two fish types (hatchery and wild), stratified by sampling location (fresh water, nearshore, and bay) and month. As we were interested in summarizing raw data rather than making inferences (explored further with statistical models below), statistical tests and pairwise comparisons for differences in prevalence between groups were not conducted. However, to visually appreciate the differences between groups, error bars (representing 95% exact binomial confidence intervals) were added to the bar graphs. The spatiotemporal patterns of the load of individual infectious agents were presented with boxplots for both fish types by sampling location and month. Additionally, summary statistics for fork length and weight of sampled fish were calculated and the distribution was shown using boxplots.
Logistic regression models for the individual infectious agents
We used logistic regression models for each individual agent, with absence or presence of the particular infectious agent (0/1) as the dependent variable, to describe the spatiotemporal variability in the prevalence of detected infectious agents. To avoid zero cell counts for some variables, models were built for infectious agents with a minimum of 5% overall prevalence. Additionally, samples collected in September 2014 were excluded from the analyses. Sampling location (FW, nearshore, and bay), month, and type of fish (wild vs. hatchery) were included a priori in the model, irrespective of their significance. To assess variation in the prevalence of infectious agents in hatchery and wild fish across the three sampling locations, an interaction term between these variables (fish type and sampling location) was included. To account for condition of the fish, a dichotomous condition factor variable, based on residuals from a model of (ln) fork length on (ln) weight, was included in the model. Positive residual values were classified as good condition factor, whereas negative residuals were considered poor (Cone 1989). To adjust for multiple comparisons among the sampling locations, fish type (or their interaction), and sampling months, Bonferroni corrections were applied. Only significant (p ≤ 0.05) spatiotemporal effects after Bonferroni corrections are reported, and are plotted separately to show the differences in predicted probabilities for (1) the sampling locations by the type of fish interaction or (2) the sampling months. For those pathogens not detected in any samples from FW locations (in both hatchery and wild fish), we built the models based on samples from the SW environment only and compared the differences between nearshore and bay locations. Similarly, for one infectious agent (P. pseudobranchicola), sampling location was not included in the model because the agent was only detected in samples from the bay.
Once the final logistic regression models describing spatiotemporal patterns for each infectious agent were identified, models were extended to identify significant associations between infectious agents using a forward stepwise procedure. For these analyses, we were only interested in the statistical significance and impact of other infectious agents on the infectious agent of interest, after adjusting for fish type, sampling location, sampling month, and condition factor. The selection of infectious agents included in the above models was based on those having unconditional associations (p < 0.2) with the infectious agent of interest; each pair of infectious agents was individually assessed using a χ2 test to determine unconditional associations.
Some hatchery fish may have been misclassified as wild fish, as 5.5% of hatchery fish did not get an adipose fin clip. We, therefore, carried out a sensitivity analysis to evaluate the potential impact this misclassification could have on our model estimates. Our data on fish fork length and weight indicated that hatchery fish were consistently larger than wild fish, with more apparent differences in the earlier months. With this knowledge, we re-labeled a portion of the larger wild fish as having been potentially misclassified. More specifically, we identified wild fish with weights in the top 10th and 20th percentiles (two separate analyses) for each location and sampling date, and re-ran the logistic models (that described the spatiotemporal variability in the prevalence of detected infectious agents) without these larger fish to compare results.
Linear regression models for the load distribution of individual infectious agents
We used linear regression models to analyze the variability in load distribution for individual infectious agents (detected at >5% overall prevalence). This analysis included only those samples classified as positive for each infectious agent. For these models, we applied algorithms similar to those used for logistic regression models, with the variability in loads described across fish type, sampling location, and month. Fork length and weight of fish were accounted for by including condition factor, and multiple test corrections were made using the Bonferroni procedure.
Models for infectious agent diversity and coexistence patterns
To evaluate the factors associated with infectious agent diversity (counts of uniquely detected infectious agents in each sampled fish), we first plotted the distribution of diversity for each sampling month by fish type (hatchery or wild) and sampling location. Then, we built a Poisson regression model with infectious agent diversity as the outcome, after evaluating for over-dispersion, with fish type, sampling location, sampling month, log fork length, and log weight as predictors.
We used cluster analysis to identify infectious agents likely to cluster or coexist. This analysis used an unsupervised hierarchical cluster analysis algorithm, available in the Pvclust package (Suzuki and Shimodaira 2006) in R version 3.4.1 (R Core Team 2015). A binary method of similarity between the dichotomous infectious agent variables and the between-group average method was applied as the dissimilarity measure between clusters, also known as the unweighted pair group method with arithmetic mean (UPGMA). After running 10 000 iterations using bootstrap resampling techniques, we identified significant clusters with approximately unbiased probability values (AU p-value) of >95% (equivalent to p < 0.05). The advantage of using the unsupervised approach is that it does not require an a priori knowledge of the optimum number of clusters (Řezanková and Everitt 2009).
Finally, we ran a log-linear model (Agresti and Kateri 2011), including only infectious agents identified as forming a significant cluster, to quantitatively confirm the coexistence of the infectious agents. For this model, we collapsed the dichotomous variables for the infectious agents into a frequency table. The log-linear model initially included only individual infectious agents to test the assumption of complete independence among them, and then included two-way, three-way, and higher-order interactions to test for joint independence, conditional independence, and homogenous association. However, as this overall model did not account for fish type or sampling location and month, we stratified the log-linear models for each of these factors and built individual log-linear models to describe the patterns of coexistence for these groups of samples.
Results
Descriptive summary
We collected 556 juvenile chinook samples from the Cowichan River system in the spring and summer of 2014. Of the 45 infectious agents included on the BioMark platform, 19 agents (5 bacterial, 2 viral, and 12 parasitic) were detected, with a prevalence range from 0.2% to 57.6%. The prevalence of individual infectious agents varied (visual trend, not based on statistical tests) by fish type, sampling location (Table 2), and across sampling months (Fig. 2). In general, the prevalence of infectious agents increased as the fish moved from FW to the bay and with sampling month. There were some exceptions to these general findings, specifically when type of fish was also considered. Ceratonova shasta was detected at a higher prevalence in FW hatchery fish than in SW fish (both hatchery and wild) and FW wild fish (Table 2). Gill chlamydia was present at a low prevalence in FW hatchery fish but was not detected in FW wild fish. Myxobolus arcticus Pugachev & Khokhlov, 1979 was detected in more than half of the wild fish in FW, but was present at a very low prevalence in FW hatchery fish. The prevalence of the FW bacterium, Flavobacterium psychrophilum Bernardet & Grimont, 1989, was higher in wild fish sampled in FW than in respective hatchery fish. Interestingly, the prevalence of this bacterium in hatchery fish increased in nearshore samples. Paranucleospora theridion, Parvicapsula kabatai Jones, Prosperi-Porta & Dawe, 2006 and P. pseudobranchicola were only present in samples from SW. For other infectious agents, prevalences were generally higher in wild fish, irrespective of sampling location. The prevalences of most other infectious agents were negligible in the FW environment, except for I. multifiliis, which was detected in >7% of wild fish sampled in FW natal sites. Viral hemorrhagic septicemia virus (VHSV) was detected in a single hatchery fish sampled in the bay.
Table 2.
| Hatchery fish (%) | Wild fish (%) | ||||||
|---|---|---|---|---|---|---|---|
| Infectious agent | Hatchery (n = 34) | Nearshore (n = 39) | Bay (n = 78) | River (n = 91) | Nearshore (n = 101) | Bay (n = 213) | Overall (%) (n = 556) |
| Myxobolus arcticus | 2.9 | 38.5 | 61.5 | 52.7 | 52.5 | 72.8 | 57.6 |
| Candidatus Branchiomonas cysticola | 2.9 | 7.7 | 67.9 | 17.6 | 62.4 | 72.8 | 52.3 |
| Paranucleospora theridion | 0.0 | 43.6 | 65.4 | 0.0 | 31.7 | 75.1 | 46.8 |
| Gill chlamydia | 2.9 | 23.1 | 47.4 | 0.0 | 40.6 | 52.8 | 36.0 |
| Parvicapsula kabatai | 0.0 | 20.5 | 11.5 | 0.0 | 7.9 | 14.6 | 10.1 |
| Parvicapsula pseudobranchicola | 0.0 | 0.0 | 15.4 | 0.0 | 0.0 | 15.5 | 8.1 |
| Ceratonova shasta | 20.6 | 0.0 | 5.1 | 5.5 | 1.0 | 8.5 | 6.3 |
| Flavobacterium psychrophilum | 2.9 | 10.3 | 2.6 | 18.7 | 2.0 | 0.9 | 5.0 |
| Parvicapsula minibicornis | 0.0 | 0.0 | 6.4 | 0.0 | 2.0 | 8.0 | 4.3 |
| Tetracapsuloides bryosalmonae | 0.0 | 0.0 | 2.6 | 0.0 | 4.0 | 8.0 | 4.1 |
| Ichthyophthirius multifiliis | 0.0 | 0.0 | 5.1 | 7.7 | 2.0 | 2.8 | 3.4 |
| Renibacterium salmoninarum | 0.0 | 0.0 | 2.6 | 0.0 | 1.0 | 7.0 | 3.2 |
| Erythrocytic necrosis virus | 0.0 | 0.0 | 6.4 | 0.0 | 0.0 | 2.8 | 2.0 |
| Ichthyophonus hoferi | 0.0 | 0.0 | 1.3 | 0.0 | 1.0 | 3.3 | 1.6 |
| Loma salmonae | 0.0 | 0.0 | 1.3 | 1.1 | 3.0 | 1.9 | 1.6 |
| Facilispora margolisi | 2.9 | 2.6 | 1.3 | 0.0 | 0.0 | 2.3 | 1.4 |
| Kudoa thyrsites | 0.0 | 0.0 | 1.3 | 0.0 | 0.0 | 2.8 | 1.3 |
| Piscichlamydia salmonis | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.4 | 0.5 |
| Viral hemorrhagic septicemia virus | 0.0 | 0.0 | 1.3 | 0.0 | 0.0 | 0.0 | 0.2 |
Fig. 2.
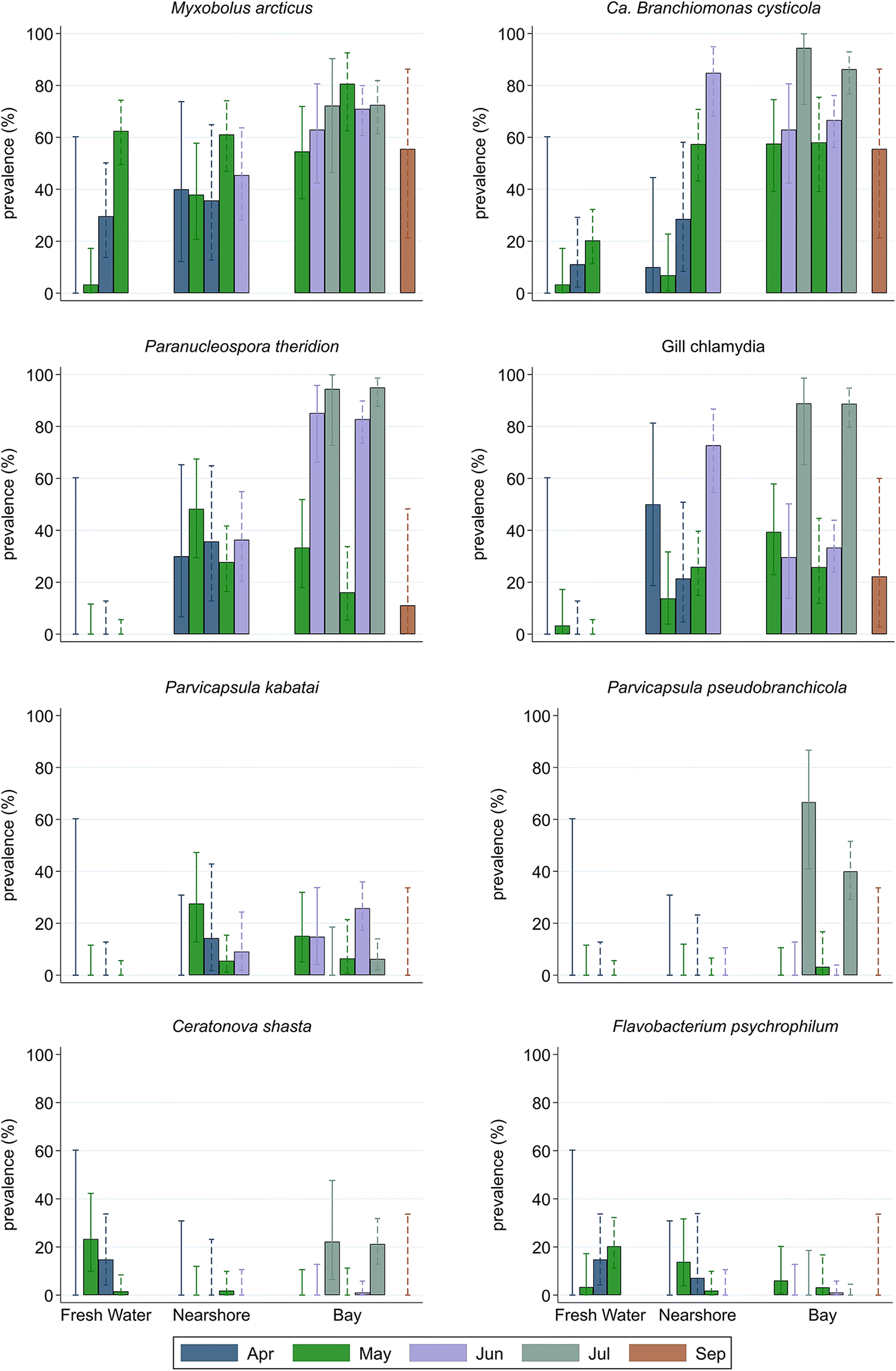
The load (copy number) of most infectious agents, similar to prevalence, increased over time (sampling month) and as fish moved from their spawning grounds in FW to the nearshore environment, and to the offshore (bay) environment. In fact, for all infectious agents except the FW bacterium F. psychrophilum, agent loads were higher in fish sampled in the bay and increased with sampling month. Although infectious agent loads varied between hatchery and wild fish across sampling locations and months (Fig. S1), no consistent visual patterns were apparent.
The mean fork length and weight of sampled fish were 78.1 mm and 6.2 g, respectively, and varied significantly. We assessed these using multiple linear regression with fork length and weight as outcomes, by type of fish (p < 0.001), sampling location (p < 0.001), and sampling month (p < 0.001). In general, hatchery fish were larger than wild fish (Figs. S2 and S3).
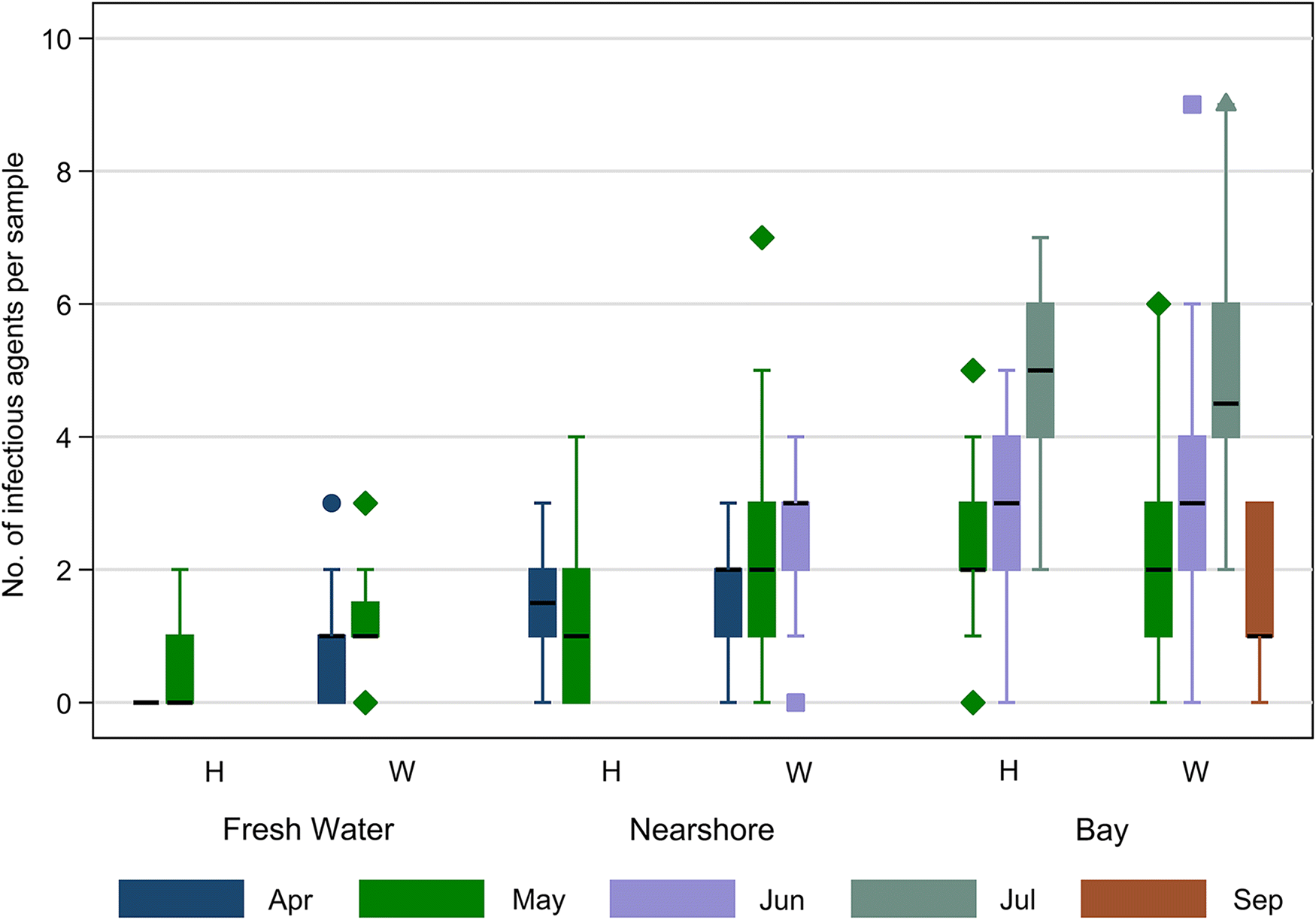
Approximately 14% (n = 77) of sampled fish did not harbor a single infectious agent, the majority of which (n = 47) were sampled in FW locations, with about 13% of the agent-free samples (n = 10) from the bay. The number of infectious agents found on a single fish ranged from 0 to 9, with a median of 2. Infectious agent diversity varied with fish type, sampling location, and month, generally increasing over time and space, with the exception of the September sample (Fig. 3).
Fig. 3.
Models for the individual infectious agents
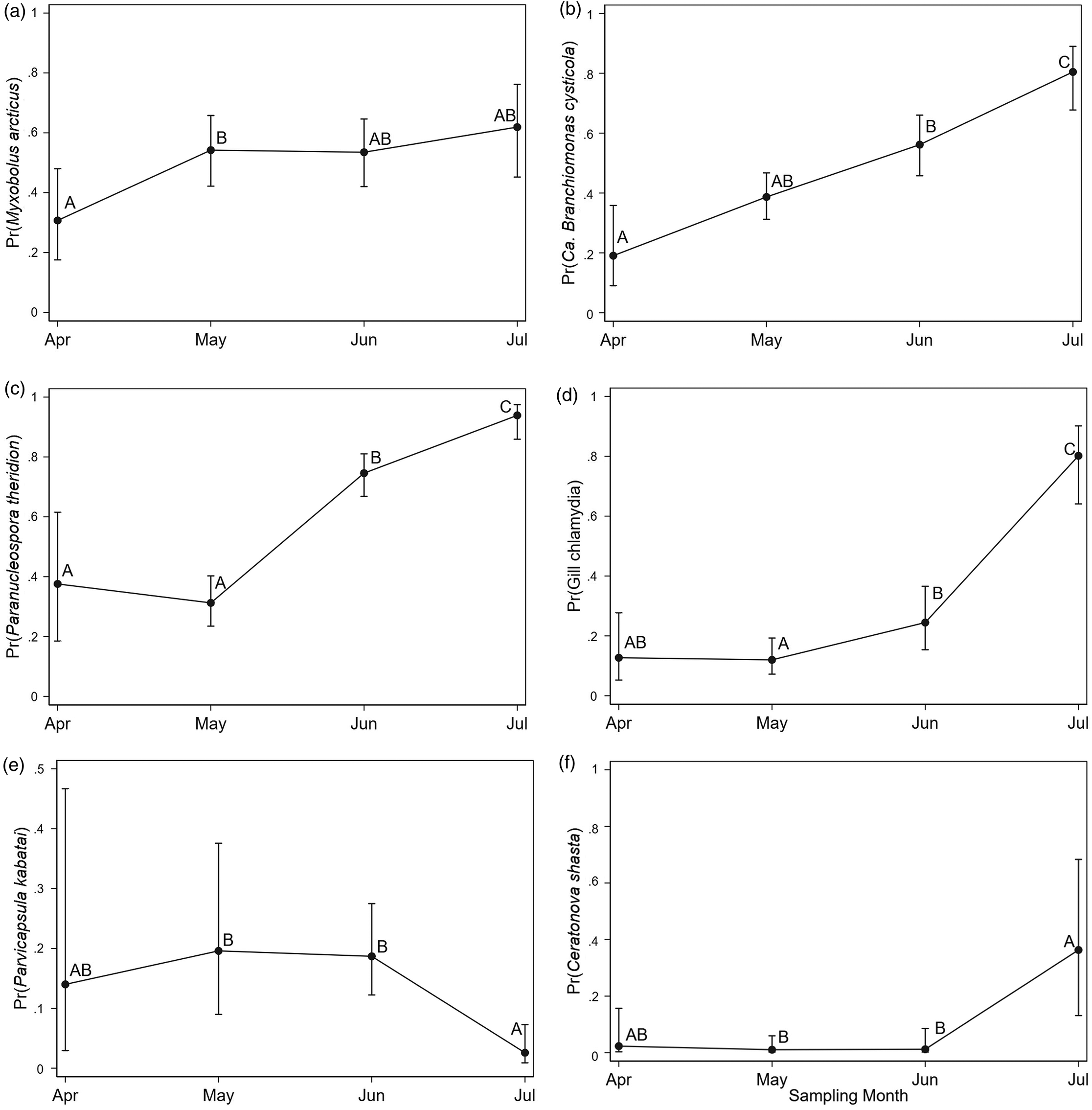
Eight infectious agents were detected at prevalence levels ≥5%, and the patterns of distribution of these agents were further explored in our models. The logistic regression models for the presence of individual infectious agents indicated that, after accounting for the effect of sampling month, prevalences of agents in hatchery and wild fish in bay samples were not significantly different (Table 3 and Figs. 4 and 5). For all infectious agents described, the trend in agent prevalence tended to converge for the two fish types once they reached the bay, irrespective of their prevalence in natal or nearshore sites. In FW, differences in the prevalence of infectious agents between hatchery and wild fish detected by our models were only significant for M. arcticus. In contrast, the effect of sampling month was significantly associated with the prevalence of six of the eight infectious agents evaluated and represented an increasing trend. For some of the infectious agents (gill chlamydia and F. psychrophilum), there were no significant differences between hatchery and wild fish, but sampling location and month were the main predictors associated with variations in prevalence. For C. shasta, the model identified a significant interaction between fish type and sampling location, but no significant differences between the groups were detected after adjusting for multiple comparisons. Condition factor was significantly associated with three infectious agents (negative association with M. arcticus and positive association with P. kabatai and P. pseudobranchicola) after accounting for spatiotemporal effect.
Table 3.
| Type#sampling locationc | Sampling location | Type (wild)d | Month | Condition factor (good) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Infectious agent | na | pb | OR | p | OR | p | OR | 95% CI | p | OR | p | OR | 95% CI | p |
| Myxobolus arcticus | 468 | <0.001 | see Fig. 4a | 0.003 | — | — | — | — | — | see Fig. 5a | 0.031 | 0.58 | 0.37–0.91 | 0.018 |
| Candidatus Branchiomonas cysticola | 547 | <0.001 | see Fig. 4b | <0.001 | — | — | — | — | — | see Fig. 5b | <0.001 | — | — | — |
| Paranucleospora theridione | 422 | <0.001 | see Fig. 4c | 0.004 | — | — | — | — | — | see Fig. 5c | <0.001 | — | — | — |
| Gill chlamydia | 546 | <0.001 | — | — | see Fig. 4d | <0.001 | 0.93 | 0.55–1.55 | 0.788 | see Fig. 5d | <0.001 | — | — | — |
| Parvicapsula kabataie | 343 | <0.001 | see Fig. 4e | 0.011 | — | — | — | — | — | see Fig. 5e | 0.001 | 2.45 | 1.19–5.03 | 0.015 |
| Parvicapsula pseudobranchicolaf | 217 | <0.001 | — | — | — | — | 0.74 | 0.32–1.73 | 0.493 | — | — | 12.10 | 4.49–32.40 | 0.001 |
| Ceratonova shasta | 508 | <0.001 | see footnote | 0.010g | — | — | — | — | — | see Fig. 5f | 0.001 | — | — | — |
| Flavobacterium psychrophilum | 449 | <0.001 | — | — | see footnote | 0.044g | 1.48 | 0.59–3.70 | 0.398 | — | 0.120 | — | — | — |
Note: Cells with no data represent terms not included in the model or not significant (for condition factor). OR, odds ratio; CI, confidence interval.
a
Total observations (n) in each model was not always equal due to missing values for the condition factor or the absence of samples from certain locations.
b
The overall p-value of the model.
c
Represents the type and sampling location interaction term.
d
For fish type, hatchery fish were used as reference category.
e
Positive samples only from nearshore and bay.
f
Positive samples only from bay, month not included in the model as it perfectly predicted the outcome.
g
Not significantly different after Bonferroni adjustment for multiple pairwise comparisons.
Fig. 4.
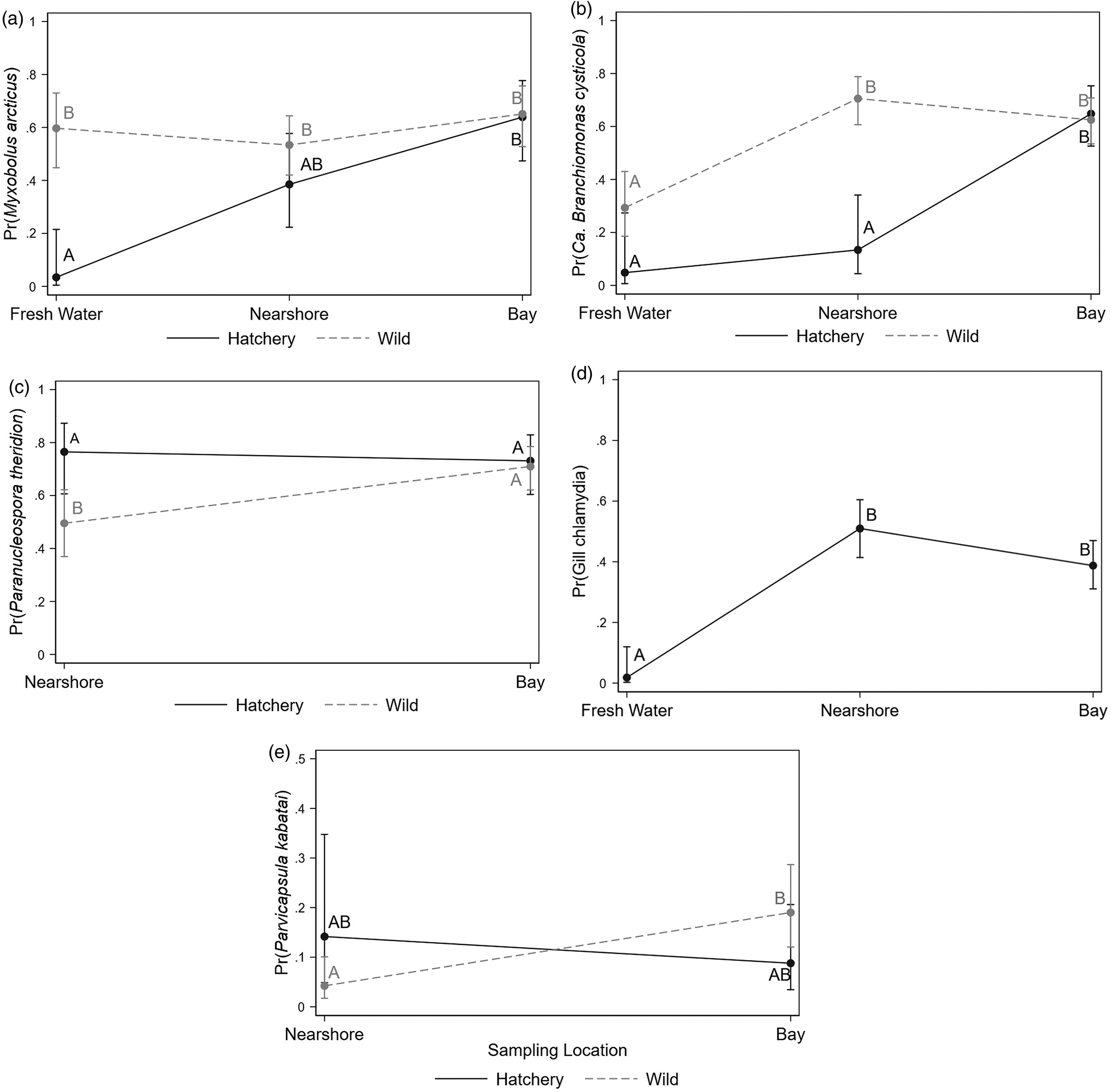
Fig. 5.
The sensitivity analysis, assessing the potential impact from misclassification of some hatchery fish as wild fish, revealed very similar spatiotemporal patterns for most (seven out of eight) of the infectious agents reported in Table 3 and Figs. 4 and 5. However, a difference was observed when evaluating P. kabatai and removing wild fish with weights in the top 20th percentile (n = 73 fish), where there was no longer a significant difference for the location and type interaction term after the Bonferroni correction (results not presented).
Some of the additional variability in the prevalence of these infectious agents, after accounting for the effects of type and condition factor of fish, sampling location, and sampling month, was explained by co-infection. Two infectious agents (P. pseudobranchicola and gill chlamydia) were associated with four different infectious agents. For example, after accounting for the above factors, Ca. B. cysticola was 2.05 times more likely to be detected in a sample that also had P. theridion, and 1.94 times more likely to be detected in a sample with gill chlamydia. The odds ratio and p-values for all the significant co-infections are presented in Table 4.
Table 4.
| Infectious agents of interest | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Co-infecting infectious | M. arcticus | Ca. B. cysticola | P. theridion | Gill chlamydia | P. kabatai | P. pseudobranchicola | C. shasta | |||||||
| agents | Odds ratio | p | Odds ratio | p | Odds ratio | p | Odds ratio | p | Odds ratio | p | Odds ratio | p | Odds ratio | p |
| Myxobolus arcticus | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Candidatus Branchiomonas cysticola | — | — | — | — | 2.02 | 0.007 | 2.11 | <0.001 | — | — | — | — | — | — |
| Paranucleospora theridion | 1.90 | 0.026 | 2.05 | 0.009 | — | — | — | — | 6.00 | <0.001 | — | — | — | — |
| Gill chlamydia | 1.81 | 0.030 | 1.94 | 0.010 | — | — | — | — | 4.10 | <0.001 | 14.2 | 0.001 | — | — |
| Parvicapsula kabatai | — | — | — | — | 5.11 | <0.001 | 2.83 | <0.001 | — | — | — | — | 6.51 | 0.020 |
| Parvicapsula pseudobranchicola | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Ceratonova shasta | — | — | — | — | — | — | — | — | 5.80 | 0.032 | — | — | — | — |
| Flavobacterium psychrophilum | — | — | — | — | — | — | — | — | — | — | a | a | — | — |
| Parvicapsula minibicornis | — | — | — | — | — | — | — | — | — | — | 2.83 | 0.050 | 4.43 | 0.020 |
| Kudoa thyrsites | — | — | — | — | — | — | — | — | 45.70 | 0.002 | b | b | 15.90 | 0.020 |
| Tetracapsuloides bryosalmonae | — | — | — | — | — | — | — | — | — | — | 7.02 | 0.002 | — | — |
| Ichthyophthirius multifiliis | — | — | 0.24 | 0.023 | — | — | — | — | — | — | — | — | — | — |
| Renibacterium salmoninarum | — | — | — | — | — | — | — | — | — | 6.54 | 0.002 | — | — | |
a
Predicts failure perfectly (when F. psychrophilum = 0, P. pseudobranchicola = 0).
b
Predicts success perfectly (when K. thyrsites = 0, P. pseudobranchicola = 1).
The load analysis for each infectious agent, using linear regression models, showed patterns similar to those observed for the presence of infectious agents (logistic regression models). For eight of the infectious agents evaluated, only the load of P. theridion was significantly different between fish types and sampling locations (higher in wild fish sampled in bay; Table S2 and Fig. S4). Interestingly, the effect of sampling month showed a significant increasing trend (Fig. S5) for four of the infectious agents (M. arcticus, Ca. B. cysticola, gill chlamydia, and P. theridion). In addition, (good) condition factor was associated with higher loads of P. theridion.
Models for infectious agent diversity and coexistence patterns
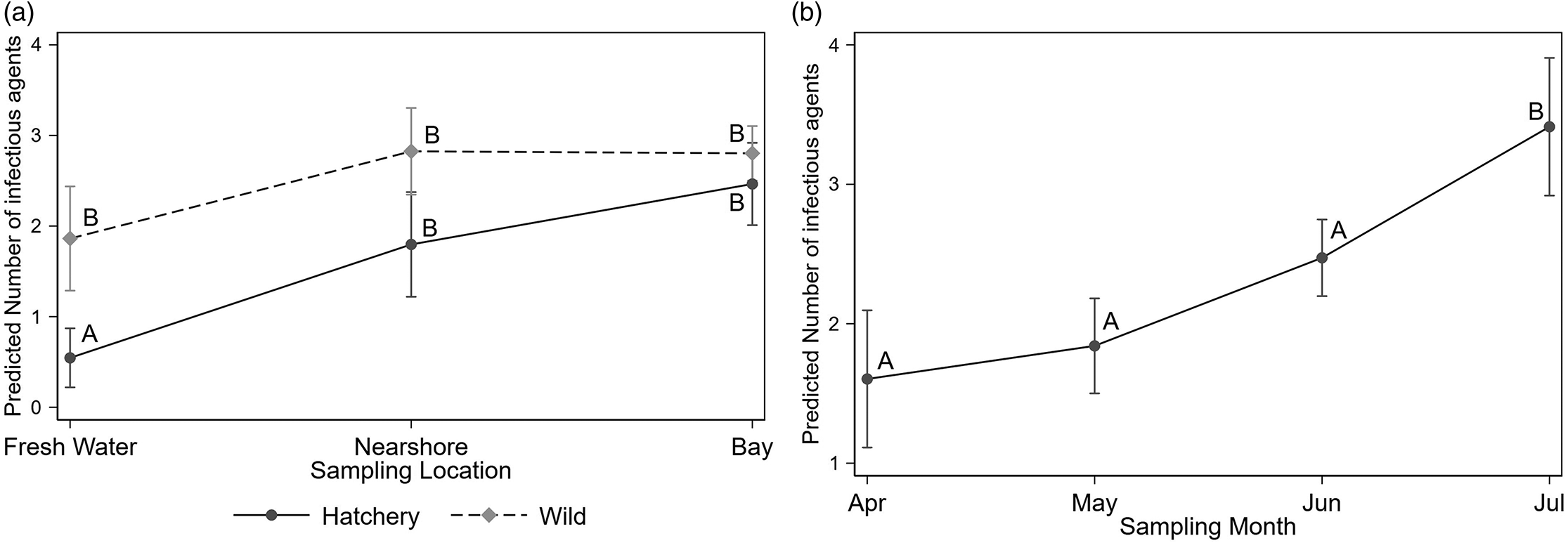
The Poisson regression model identified that fork length, an interaction between fish type and location (p < 0.001), and the sampling month (p < 0.001) were all significantly associated with infectious agent diversity. The model estimated, for example, that an increase in fork length from 61 to 92 mm (representing the 25th and 75th percentiles in our study population) increased the number of infectious agents by 1.33 (95%CI 1.10–1.56). Infectious agent diversity was significantly lower in hatchery fish sampled from FW, but once they moved to nearshore or bay sites diversity converged between the two fish types (Fig. 6a). Similarly, infectious agent diversity increased with sampling month (Fig. 6b).
Fig. 6.
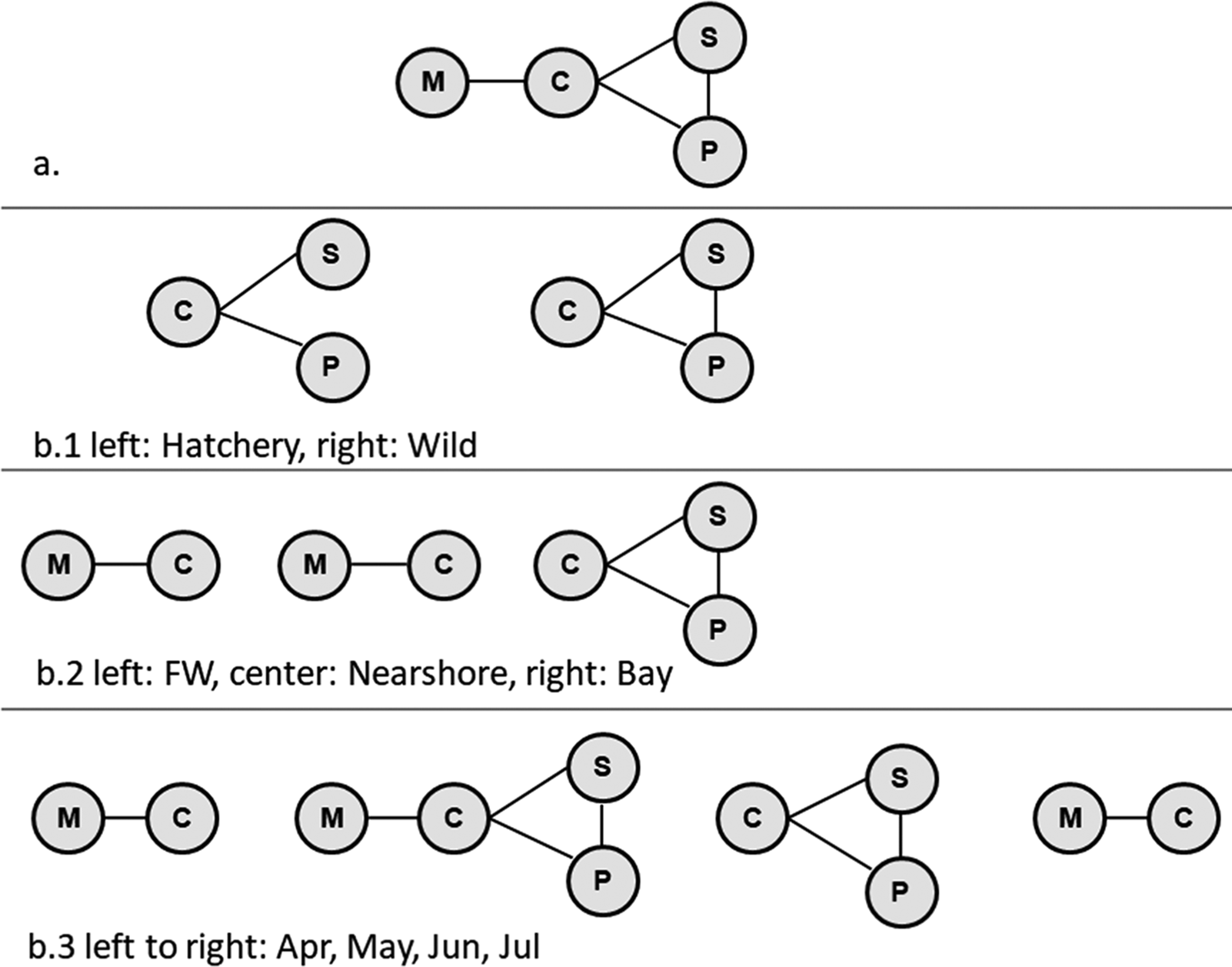
Cluster analysis of the infectious agents identified two significant clusters. The first cluster consisted of M. arcticus, Ca. B. cysticola, gill chlamydia, and P. theridion, whereas the second included Loma salmonae (Putz, Hoffman & Dunbar, 1965), and Piscichlamydia salmonis Draghi, Popov, Kahl, Stanton, Brown, Tsongalis, West & Frasca 2004 (Fig. 7). Further exploration of the relationships among the members of the first significant cluster, using log-linear models, without consideration of fish type, sampling location, or month, showed that M. arcticus was associated with Ca. B. cysticola, but conditionally independent of P. theridion and gill chlamydia. Alternately, Ca. B. cysticola, P. theridion, and gill chlamydia were interdependent or positively associated with each other (Fig. 8a). When we stratified this analysis for each fish type, sampling location, and sampling month, significant two- and three-way interactions among these infectious agents were apparent (Fig. 8b). The three-way interaction (interdependence) among Ca. B. cysticola, P. theridion, and gill chlamydia was consistently present in wild fish (without considering location and sampling month), in samples from the bay, and in samples collected in May and June. Similarly, a separate log-linear model for infectious agents from the second cluster suggested that L. salmonae and P. salmonis were highly associated in wild fish for samples collected from the bay and in June (p < 0.05, results not shown).
Fig. 7.
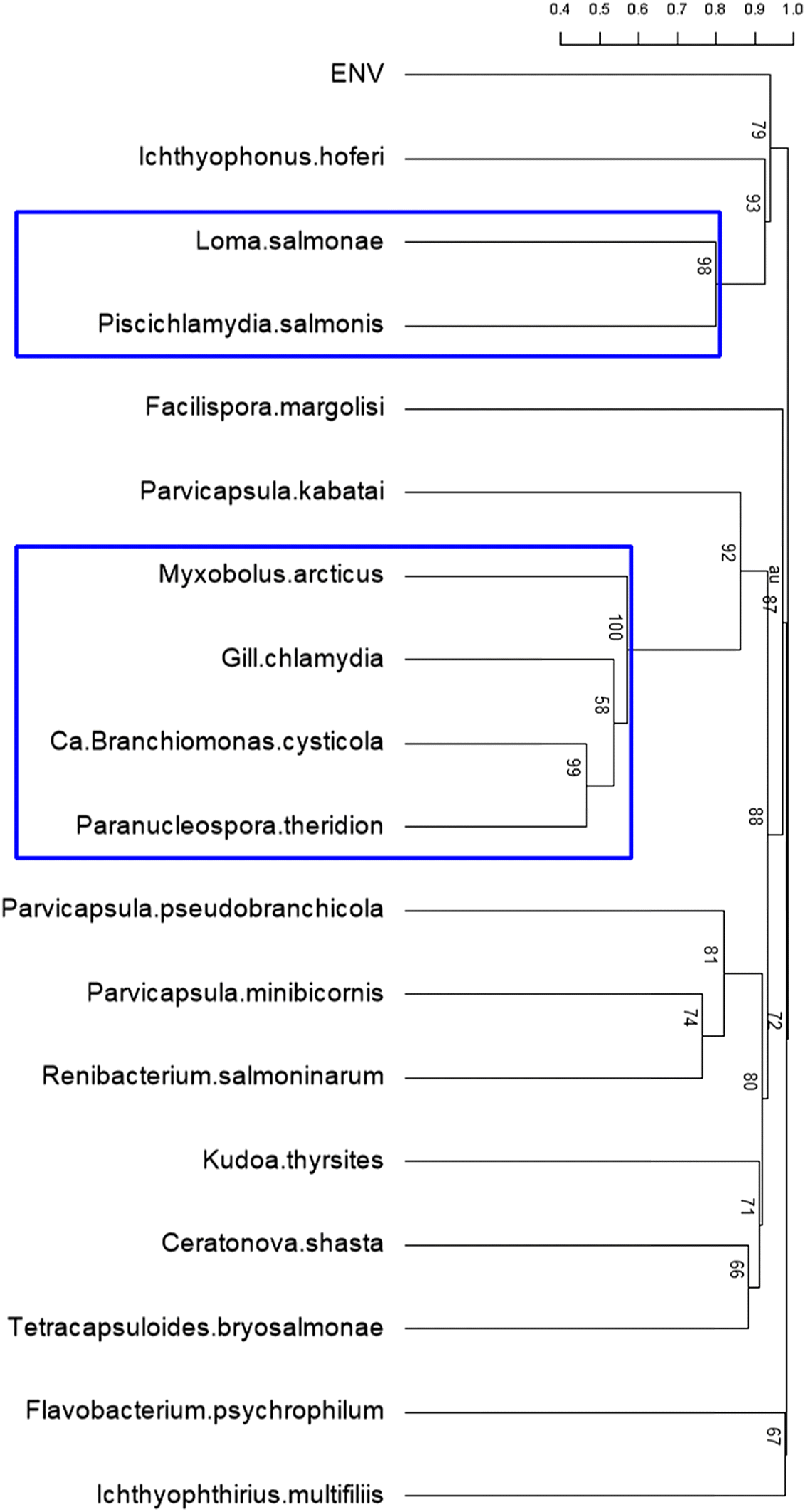
Fig. 8.
Discussion
Our study provides insights into the prevalence and distribution of dozens of infectious agents in hatchery-reared and wild-caught juvenile chinook salmon from the Cowichan River watershed in BC Canada. We found that in FW rearing environments, hatchery and wild salmon showed different trends in prevalence, load, and diversity for many of FW agents, likely reflecting differential exposures to agents in these environments. Interestingly, agent diversity was higher in wild chinook salmon. However, most agent profiles converged in hatchery and wild fish cohabitating in the nearshore and bay marine environments. These data suggest that the environment in which fish are sampled has a stronger effect on agent profiles than their origin as cultured or wild fish, as both show susceptibility to infection by a range of infectious agents. Our data fall short, however, of documenting whether these agents are causing disease, or whether hatchery and wild fish differ in their susceptibility to disease.
The prevalence of most infectious agents was significantly associated with sampling month, and was higher during summer than spring. This relationship may be a manifestation of elevated water temperatures (which range from 11–12 °C in May to 15–17 °C in July) enhancing the activity of infectious agents or merely the temporal nature of accumulation and propagation of infectious agents through fish populations. There was some evidence that the temporal effect varied for the two fish types, as it appears in the descriptive statistics, but their interactions were not evaluated because of the limited sample size. The increase in infectious agent loads between sampling months could, similarly, be associated either with replication activity in elevated water temperatures or accumulations over time. Most infectious agents peaked in prevalence and load in the bay, where we hypothesize that juveniles would be exposed to higher abundances of marine fish that may be carriers of agents that can infect salmon.
The diversity of infectious agents, which has previously been shown to be associated with risk of predation (Miller et al. 2014), was higher in Cowichan juvenile chinook salmon in our study than in previously published reports for the region (Miller et al. 2014; Bass et al. 2017), and may represent an increased predation risk for this population. However, Miller et al. (2014) did not include all the infectious agents incorporated in this study, and Bass et al. (2017) evaluated infectious agents in returning adult chinook salmon only.
Agent diversity in juvenile Cowichan River chinook salmon increased with the size of the fish. As fish size was also associated with sampling location, thereby confounding the association between sampling location and infectious agent diversity (>20% change in the coefficient of fish type and sampling location interaction in the Poisson regression model), our data suggest that larger fish were more likely to reach the bay and be exposed to a diverse range of pathogens, whereas smaller fish initially stayed near shore before venturing further into the bay.
Hatchery fish were generally larger than wild fish during any given sampling event; in fact, in early sample dates the weight distributions between hatchery and wild fish were virtually non-overlapping. Given this finding, we were able to use weight of fish to assess the potential impact that misclassification of hatchery fish as wild fish could have had on our results. This sensitivity analysis suggests that misclassification had little impact on our results.
Multiple years of data from the Cowichan system show that hatchery fish are observed in the nearshore environment within day(s) of release from the hatchery, indicating rapid movement through freshwater. These fish are also observed in the deeper water of the bay within days of ocean entry, suggesting a short residency in the nearshore environment. In contrast, smaller wild chinook salmon are observed in the freshwater and nearshore environments over an extended period of time (Chittenden et al. 2018). It is important to note that our experimental design was aimed at balancing sample sizes for hatchery and wild fish within environments over time, which did not necessarily reflect their actual proportions (i.e., hatchery and wild fish relative abundance) at each sampling location and month. Hence, although we show that hatchery and wild fish may both be vulnerable to becoming infected in each of these environments, they may not necessarily become infected at the same rates. For example, we show that agent diversity and load increase as fish move from the nearshore to the offshore environment. Over multiple years, hatchery fish have been observed in offshore waters weeks before wild fish are detected (Chittenden et al. 2018). From this observation, we surmise that if hatchery fish move offshore more quickly than wild fish they would likely experience higher agent diversity and load soon after ocean entry, when they are still adjusting to the environmental changes that accompany the transition from FW to SW, and may be more vulnerable to additional stressors. In the future, it will be important to demonstrate the actual time that hatchery and wild fish spend in each environment, ideally with tracking data, although there may be limitations (Tiffan et al. 2015) on tagging smaller wild fish (<40 mm).
Our findings are based solely on the detection of the nucleic acids of the infectious agents in the sampled fish, which does not necessarily imply the presence of associated infectious diseases in the studied population. However, the molecular presence of an agent can be, in some scenarios, a risk factor associated with the development of clinical disease. In addition to agents, environmental factors such as water quality, oxygen level, water temperature, toxins, and fish condition (physical fitness and presence of other disease or co-infections) may lead to increased risk of pathological changes, morbidity, and mortality in a population (Salonius and Iwama 1993; Castro et al. 2013). Combining findings from epidemiological and histopathological studies, with gene expression profiling of host immune genes to assess molecular signatures of disease (e.g., see (Miller et al. 2017), may allow us to unravel the linkages between infectious agents, host susceptibility, and the likelihood of disease, and will enhance our understanding of the impacts of infectious agents/diseases on Pacific salmon populations.
Four of the eight infectious agents detected with >5% overall prevalence in our study (M. arcticus, C. shasta, P. kabatai, and F. psychrophilum) are known endemics of BC (Kent 2011; Miller et al. 2014) and are relatively well studied (P. kabatai is the exception). The remaining four agents (P. theridion, gill chlamydia, P. pseudobranchicola, and Ca. B. cysticola) have only recently been identified in BC salmon, most by other studies in our program (Miller et al. 2014; Bass et al. 2017), and we are just beginning to develop an understanding of their distributions and potential for impact in Pacific salmon. We discuss the implications of the detected infectious agents in the supplementary material.
It is important to emphasize that although we detected 19 distinct infectious agents in the out-migrating Cowichan juvenile chinook, some previously associated with large-scale mortality events (C. shasta, and P. minibicornis) and some only recently reported or known (see above), this study also provides information about a number of agents not observed across our sample of 556 chinook salmon. Most notably, three viruses reportable to the World Organisation for Animal Health (OIE) (infectious salmon anemia virus, infectious pancreatic necrosis virus, and salmonid alphavirus) capable of causing devastating impacts in cultured fish were not observed, nor was the endemic infectious hematopoietic necrosis virus (IHNV). Moreover, the Piscine orthoreovirus (PRV) Rimstad and Wessel, 2015, which is associated with an emerging heart disease in Norway and was recently identified in farmed Atlantic salmon in BC (Di Cicco et al. 2017), was not detected in Cowichan chinook salmon. Our sample sizes were not sufficient to establish a “freedom from infection” status (Cameron and Baldock 1998), which was beyond the scope of this study, but our findings indicate that these agents were not present above a 1% prevalence level, with a confidence level of 95%, assuming a test with 95% sensitivity and specificity.
Findings from the log-linear models indicated that the infectious agents previously associated with gill diseases, namely Ca. B. cysticola, P. theridion, and gill chlamydia (Steinum et al. 2010; Mitchell et al. 2013; Gunnarsson et al. 2017), were observed as co-infections more often than expected by chance. The co-existence of these three pathogens in the Cowichan chinook stock reinforces previous observations of their co-occurrence, but raises questions about whether it could lead to clinical observations similar to those seen in farmed Atlantic salmon in Norway (Steinum et al. 2010; Mitchell et al. 2013; Gunnarsson et al. 2017). In future, histopathology could clarify the association, if any, of these agents with gill disease in BC salmon.
It is worth noting some limitations of the current study and the study design. As we did not follow the same fish over time and space, the spatiotemporal trends of infectious agent prevalence represent the group level effect and may not be true for individual fish. Similarly, we may have underestimated the prevalence of agents causing acute diseases, such as C. shasta, VHSV, and IHNV, as they may have caused mortalities in infected fish before sampling; the monthly temporal frame of sampling may not have been appropriate to detect such shifts in prevalence.
This study provides new information on the spatiotemporal distribution and prevalence of infectious agents in one stock of hatchery-reared and wild-caught juvenile chinook salmon on southern Vancouver Island. Although the study focused on the short time period during the transition from FW to SW environments, a period thought to be one of the most challenging for salmon survival, our findings indicated that wild and hatchery fish were most divergent in agent profiles in their natal sites. Differences in prevalence dissipated once they converged in the marine environment, suggesting that hatchery and wild fish are susceptible to a similar array of infectious agents when in a common environment. However, given that hatchery fish are larger than wild fish, and as these larger fish appear to be moving into the deeper waters of the bay earlier than wild fish, we hypothesize that hatchery fish may experience earlier exposure to a diversity of agents post SW entry than do wild fish. The use of a similar approach for other stocks and species could expand our knowledge of the temporal and spatial variance in agent profiles in the early marine environment, and beyond. Our group intends to undertake similar studies for other genetically and geographically distinct chinook, coho, and sockeye salmon stocks from several river systems in BC to gain a broader understanding of distribution and prevalence of infectious agents and potentially elucidate the relative contribution of infectious disease(s) in salmon declines.
Acknowledgements
This work was funded by Genome British Columbia, Pacific Salmon Foundation and the Mitacs project (IT06621) under phase 2b of the Strategic Salmon Health Initiative and the Salish Sea Marine Survival Program, and partly funded by the Canada Excellence Research Chair in Aquatic Epidemiology. We thank the Cowichan hatchery for allowing access to fish for sampling, and co-op students (Dylan Conover, Katarina Wind, and Breanne Urquhart) and staff (Karia Kaukinen and Norma Ginther) involved in sampling and initial sample processing in the molecular genetics laboratory at the Pacific Biological Station. We are thankful to Henrik Stryhn for insightful discussions on some of the statistical methods applied in this manuscript. We also wish to thank William Chalmers for editorial assistance with the manuscript.
References
Agresti A, and Kateri M. 2011. Categorical data analysis. John Wiley & Sons, New York City, New York. pp. 314–356.
Araki H, Berejikian BA, Ford MJ, and Blouin MS. 2008. Fitness of hatchery-reared salmonids in the wild. Evolutionary Applications, 1(2): 342–355.
Araki H, Cooper B, and Blouin MS. 2007. Genetic effects of captive breeding cause a rapid, cumulative fitness decline in the wild. Science, 318(5847): 100–103.
Arkoosh MR, Clemons E, Kagley AN, Stafford C, Glass AC, Jacobson K, et al. 2004. Survey of pathogens in juvenile salmon Oncorhynchus Spp. migrating through Pacific Northwest estuaries. Journal of Aquatic Animal Health, 16(4): 186–196.
Bams RA. 1967. Differences in performance of naturally and artificially propagated sockeye salmon migrant fry, as measured with swimming and predation tests. Journal of the Fisheries Board of Canada, 24(5): 1117–1153.
Bass A, Hinch S, Teffer A, Patterson D, and Miller K. 2017. A survey of microparasites present in adult migrating Chinook salmon (Oncorhynchus tshawytscha) in south-western British Columbia determined by high-throughput quantitative polymerase chain reaction. Journal of Fish Diseases, 40(4): 453–477.
Beacham TD, Candy JR, Jonsen KL, Supernault J, Wetklo M, Deng L, et al. 2006. Estimation of stock composition and individual identification of Chinook salmon across the Pacific Rim by use of microsatellite variation. Transactions of the American Fisheries Society, 135(4): 861–888.
Beamish RJ, and Mahnken C. 2001. A critical size and period hypothesis to explain natural regulation of salmon abundance and the linkage to climate and climate change. Progress in Oceanography, 49(1–4): 423–437.
Beamish RJ, Riddell BE, Neville CEM, Thomson BL, and Zhang Z. 1995. Declines in chinook salmon catches in the Strait of Georgia in relation to shifts in the marine environment. Fisheries Oceanography, 4(3): 243–256.
Beamish RJ, McCaughran D, King JR, Sweeting RM, and Mcfarlane GA. 2000. Estimating the abundance of juvenile coho salmon in the Strait of Georgia by means of surface trawls. North American Journal of Fisheries Management, 20(2): 369–375.
Beamish RJ, Neville C, Sweeting R, and Lange K. 2012a. The synchronous failure of juvenile Pacific salmon and herring production in the Strait of Georgia in 2007 and the poor return of sockeye salmon to the Fraser River in 2009. Marine and Coastal Fisheries, 4(1): 403–414.
Beamish RJ, Sweeting RM, Neville CM, Lange KL, Beacham TD, and Preikshot D. 2012b. Wild chinook salmon survive better than hatchery salmon in a period of poor production. Environmental Biology of Fishes, 94(1): 135–148.
Bradford MJ, Lovy J, Patterson DA, Speare DJ, Bennett WR, Stobbart AR, et al. 2010. Parvicapsula minibicornis infections in gill and kidney and the premature mortality of adult sockeye salmon (Oncorhynchus nerka) from Cultus Lake, British Columbia. Canadian Journal of Fisheries and Aquatic Sciences, 67(4): 673–683.
Cameron AR, and Baldock FC. 1998. A new probability formula for surveys to substantiate freedom from disease. Preventive Veterinary Medicine, 34(1): 1–17.
Castro V, Grisdale-Helland B, Jørgensen SM, Helgerud J, Claireaux G, Farrell AP, et al. 2013. Disease resistance is related to inherent swimming performance in Atlantic salmon. BMC Physiology, 13(1): 1.
Cederholm CJ, Kunze MD, Murota T, and Sibatani A. 1999. Pacific salmon carcasses: essential contributions of nutrients and energy for aquatic and terrestrial ecosystems. Fisheries, 24(10): 6–15.
Chittenden CM, Sweeting R, Neville CM, Young K, Galbraith KM, Carmack E, et al. 2018. Estuarine and marine diets of out-migrating Chinook Salmon smolts in relation to local zooplankton populations, including harmful blooms. Estuarine, Coastal and Shelf Science, 200: 335–348.
Christie MR, Marine M, French R, Waples RS, and Blouin M. 2012. Effective size of a wild salmonid population is greatly reduced by hatchery supplementation. Heredity, 109(4): 254–260.
Cohen BI. 2012. The Uncertain Future of Fraser River Sockeye: The Sockeye Fishery. Final Report, Cohen Commission of Inquiry into the Decline of Sockeye Salmon in the Fraser River (Canada) [online]: Available from publications.gc.ca/collections/collection_2012/bcp-pco/CP32-93-2012-1-eng.pdf.
Cone RS. 1989. The need to reconsider the use of condition indices in fishery science. Transactions of the American Fisheries Society, 118(5): 510–514.
Di Cicco E, Ferguson HW, Schulze AD, Kaukinen KH, Li S, Vanderstichel R, et al. 2017. Heart and skeletal muscle inflammation (HSMI) disease diagnosed on a British Columbia salmon farm through a longitudinal farm study. PLoS ONE, 12(2): e0171471.
Emmerich R. 1894. Über eine durch Bacterien erzeugte Seuche unter den Forellen. Arch for Hygiene und Bakteriologie, 21: 1–21.
Fujiwara M, Mohr MS, Greenberg A, Foott JS, and Bartholomew JL. 2011. Effects of ceratomyxosis on population dynamics of Klamath fall-run Chinook salmon. Transactions of the American Fisheries Society, 140(5): 1380–1391.
Gende SM, Edwards RT, Willson MF, and Wipfli MS. 2002. Pacific salmon in Aquatic and Terrestrial Ecosystems: Pacific salmon subsidize freshwater and terrestrial ecosystems through several pathways, which generates unique management and conservation issues but also provides valuable research opportunities. Bioscience, 52(10): 917–928.
Gresh T, Lichatowich J, and Schoonmaker P. 2000. An estimation of historic and current levels of salmon production in the Northeast Pacific ecosystem: evidence of a nutrient deficit in the freshwater systems of the Pacific Northwest. Fisheries, 25(1): 15–21.
Gunnarsson G, Karlsbakk E, Blindheim S, Plarre H, Imsland A, Handeland S, et al. 2017. Temporal changes in infections with some pathogens associated with gill disease in farmed Atlantic salmon (Salmo salar L). Aquaculture, 468: 126–134.
Hallett SL, Ray RA, Hurst CN, Holt RA, Buckles GR, Atkinson SD, et al. 2012. Density of the waterborne parasite Ceratomyxa shasta and its biological effects on salmon. Applied and Environmental Microbiology, 78(10): 3724–3731.
Hedrick RP. 1998. Relationships of the host, pathogen, and environment: implications for diseases of cultured and wild fish populations. Journal of Aquatic Animal Health, 10(2): 107–111.
Hilborn R, and Winton J. 1993. Learning to enhance salmon production: lessons from the Salmonid Enhancement Program. Canadian Journal of Fisheries and Aquatic Sciences, 50(9): 2043–2056.
Hoar WS, Randall DJ, Iwama G, and Nakanishi T. 1997. The fish immune system: organism, pathogen, and environment. Academic Press, San Diego, California.
Jeffries KM, Hinch SG, Gale MK, Clark TD, Lotto AG, Casselman MT, et al. 2014. Immune response genes and pathogen presence predict migration survival in wild salmon smolts. Molecular Ecology, 23(23): 5803–5815.
Jones S, and Dawe S. 2002. Ichthyophonus hoferi Plehn & Mulsow in British Columbia stocks of Pacific herring, Clupea pallasi Valenciennes, and its infectivity to chinook salmon, Oncorhynchus tshawytscha (Walbaum). Journal of Fish Diseases, 25(7): 415–421.
Kent M. 2011. Infectious diseases and potential impacts on survival of Fraser River sockeye salmon. Cohen Commission Technical Report 1 [online]: Available from watershed-watch.org/wordpress/wp-content/uploads/2011/08/Exh-1449-CCI001131.pdf.
Kent ML, Traxler GS, Kieser D, Richard J, Dawe SC, Shaw RW, et al. 1998. Survey of salmonid pathogens in ocean-caught fishes in British Columbia, Canada. Journal of Aquatic Animal Health, 10(2): 211–219.
Kocan R, Hershberger P, and Winton J. 2004. Ichthyophoniasis: an emerging disease of Chinook salmon in the Yukon River. Journal of Aquatic Animal Health, 16(2): 58–72.
Lichatowich J, and Lichatowich JA. 2001. Salmon without rivers: a history of the Pacific salmon crisis. Island Press, Washington, D.C.
Miller KM, Li S, Kaukinen KH, Ginther N, Hammill E, Curtis JM, et al. 2011. Genomic signatures predict migration and spawning failure in wild Canadian salmon. Science, 331(6014): 214–217.
Miller KM, Teffer A, Tucker S, Li S, Schulze AD, Trudel M, et al. 2014. Infectious disease, shifting climates, and opportunistic predators: cumulative factors potentially impacting wild salmon declines. Evolutionary Applications, 7(7): 812–855.
Miller KM, Gardner IA, Vanderstichel R, Burnley T, Schulze AD, Li S, et al. 2016. Report on the performance evaluation of the Fluidigm BioMark Platform for high-throughput microbe monitoring in salmon [online]: Available from publications.gc.ca/collections/collection_2016/mpo-dfo/Fs70-5-2016-038-eng.pdf.
Miller KM, Günther OP, Li S, Kaukinen KH, and Ming TJ. 2017. Molecular indices of viral disease development in wild migrating salmon. Conservation Physiology, 5(1): cox036.
Mitchell SO, Steinum TM, Toenshoff ER, Kvellestad A, Falk K, Horn M, et al. 2013. ‘Candidatus Branchiomonas cysticola’ is a common agent of epitheliocysts in seawater-farmed Atlantic salmon Salmo salar in Norway and Ireland. Diseases of Aquatic Organisms, 103(1): 35–43.
Naish KA, Taylor JE, Levin PS, Quinn TP, Winton JR, Huppert D, et al. 2007. An evaluation of the effects of conservation and fishery enhancement hatcheries on wild populations of salmon. Advances in Marine Biology, 53: 61–194.
National Research Council. 1996. Upstream: salmon and society in the Pacific Northwest. National Academies Press, Washington, D.C.
Nehring RB, and Thompson K. 2001. Impact assessment of some physical and biological factors in the whirling disease epizootic among wild trout in Colorado. Special Report Number 76. Colorado Division of Wildlife, Fort Collins, Colorado.
Pearcy WG. 1992. Ocean ecology of North Pacific salmonids. University of Washington Press, Seattle, Washington. 178 p.
Peterman RM, and Dorner B. 2012. A widespread decrease in productivity of sockeye salmon (Oncorhynchus nerka) populations in western North America. Canadian Journal of Fisheries and Aquatic Sciences, 69(8): 1255–1260.
R Core Team. 2015. R: a language and environment for statistical computing. R foundation for Statistical Computing, Vienna, Austria. [online]: Available from r-project.org/.
Řezanková H, and Everitt B. 2009. Cluster analysis and categorical data. Statistika, 46: 216–232.
Riddell B, Bradford M, Carmichael R, Hankin D, Peterman R, and Wertheimer A. 2013. Assessment of status and factors for decline of southern BC chinook salmon: Independent panel’s report. Prepared with the assistance of DR Marmorek and AW Hall, ESSA Technologies Ltd., Vancouver, British Columbia for Fisheries and Oceans Canada, Vancouver, British Columbia and Fraser River Aboriginal Fisheries Secretariat, Merritt, British Columbia.
Salonius K, and Iwama GK. 1993. Effects of early rearing environment on stress response, immune function, and disease resistance in juvenile coho (Oncorhynchus kisutch) and chinook salmon (O. tshawytscha). Canadian Journal of Fisheries and Aquatic Sciences, 50(4): 759–766.
Sandell TA. 2010. The effects of single and multiple pathogen and parasite infections on juvenile Chinook and coho salmon during early marine residency. Ph.D. dissertation, Oregon State University, Eugene, Oregon.
Steinum T, Kvellestad A, Colquhoun D, Heum M, Mohammad S, Grøntvedt RN, et al. 2010. Microbial and pathological findings in farmed Atlantic salmon Salmo salar with proliferative gill inflammation. Diseases of Aquatic Organisms, 91(3): 201–211.
Stephen C, Stitt T, Dawson-Coates J, and Mccarthy A. 2011. Assessment of the potential effects of diseases present in salmonid enhancement facilities on Fraser River sockeye salmon. Cohen Commission Technical Report 1A [online]: Available from watershed-watch.org/wordpress/wp-content/uploads/2011/08/Exh-1454-NonRT.pdf.
Suzuki R, and Shimodaira H. 2006. Pvclust: an R package for assessing the uncertainty in hierarchical clustering. Bioinformatics, 22(12): 1540–1542.
Sweeting R, Beamish R, Noakes D, and Neville C. 2003. Replacement of wild coho salmon by hatchery-reared coho salmon in the Strait of Georgia over the past three decades. North American Journal of Fisheries Management, 23(2): 492–502.
Tiffan KF, Perry RW, Connor WP, Mullins FL, Rabe CD, and Nelson DD. 2015. Survival, growth, and tag retention in age-0 Chinook salmon implanted with 8-, 9-, and 12-mm PIT tags. North American Journal of Fisheries Management, 35(4): 845–852.
Traxler GS, Richard J, and McDonald TE. 1998. Ichthyophthirius multifiliis (Ich) epizootics in spawning sockeye salmon in British Columbia, Canada. Journal of Aquatic Animal Health, 10(2): 143–151.
Van Gaest AL, Dietrich JP, Thompson DE, Boylen DA, Strickland SA, Collier TK, et al. 2011. Survey of pathogens in hatchery Chinook salmon with different out-migration histories through the Snake and Columbia rivers. Journal of Aquatic Animal Health, 23(2): 62–77.
Weber ED, and Fausch KD. 2003. Interactions between hatchery and wild salmonids in streams: differences in biology and evidence for competition. Canadian Journal of Fisheries and Aquatic Sciences, 60(8): 1018–1036.
Supplementary Material
Supplementary Material 1 (DOCX / 1.16 MB)
- Download
- 1.16 MB
Information & Authors
Information
Published In

FACETS
Volume 3 • Number 1 • October 2018
Pages: 695 - 721
Editor: Steven J. Cooke
History
Received: 28 September 2017
Accepted: 26 March 2018
Version of record online: 5 July 2018
Copyright
© 2018 Thakur et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper and in the Supplementary Material.
Key Words
Sections
Subjects
Authors
Author Contributions
KKT, RV, and KMM conceived and designed the study.
SL, CN, and AT performed the experiments/collected the data.
KKT, RV, EL, ST, and KMM analyzed and interpreted the data.
RV and KMM contributed resources.
KKT, RV, SL, EL, ST, CN, and KMM drafted or revised the manuscript.
Competing Interests
KMM is currently serving as a Subject Editor for FACETS, but was not involved in review or editorial decisions regarding this manuscript.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Krishna K. Thakur, Raphaël Vanderstichel, Shaorong Li, Emilie Laurin, Strahan Tucker, Chrys Neville, Amy Tabata, and Kristina M. Miller. 2018. A comparison of infectious agents between hatchery-enhanced and wild out-migrating juvenile chinook salmon (Oncorhynchus tshawytscha) from Cowichan River, British Columbia. FACETS.
3(1): 695-721. https://doi.org/10.1139/facets-2017-0113
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. The spatial distribution of infectious agents in wild Pacific salmon along the British Columbia coast
2. Piscine orthoreovirus Genotype-1 (PRV-1) in Wild Pacific Salmon of British Columbia, Canada: 2011–2020
3. Expansion of the Beta-Proteobacterial Genus Ca. Ichthyocystis: A Case Report of Epitheliocystis in the Pompano Trachinotus ovatus
4. Identification of infectious agents in early marine Chinook and Coho salmon associated with cohort survival
5. Host‐pathogen‐environment interactions predict survival outcomes of adult sockeye salmon (
Oncorhynchus nerka
) released from fisheries
6. Descriptive multi-agent epidemiology via molecular screening on Atlantic salmon farms in the northeast Pacific Ocean
7. Wild salmonids are running the gauntlet of pathogens and climate as fish farms expand northwards
8. Review of infectious agent occurrence in wild salmonids in British Columbia, Canada
9. A molecular assessment of infectious agents carried by Atlantic salmon at sea and in three eastern Canadian rivers, including aquaculture escapees and North American and European origin wild stocks
10. Applying a gene-suite approach to examine the physiological status of wild-caught walleye (
Sander vitreus
)
11. Comparison of infectious agents detected from hatchery and wild juvenile Coho salmon in British Columbia, 2008-2018
12. Distribution and Phylogeny of Erythrocytic Necrosis Virus (ENV) in Salmon Suggests Marine Origin
13. Infectious agent detections in archived Sockeye salmon (
Oncorhynchus nerka
) samples from British Columbia, Canada (1985–94)
14. Histopathological and novel high-throughput molecular monitoring data from farmed salmon (Salmo salar and Oncorhynchus spp.) in British Columbia, Canada, from 2011–2013
15. Detection and Assessment of the Distribution of Infectious Agents in Juvenile Fraser River Sockeye Salmon, Canada, in 2012 and 2013