Channel geomorphology differences between stream reaches with grass- or tree-dominated riparian vegetation in southern Manitoba
Abstract
Removal of trees for pasture or crop production is common along the stream reaches in the Canadian Prairies, resulting in a patchwork of forested and nonforest riparian vegetation along most streams. The effect of vegetation type on channel geomorphology and potential to influence sediment dynamics was studied using eight paired reaches (forested and nonforest) within agricultural watersheds in southern Manitoba, Canada. High potential for bank erosion was observed at all sites (bank erosion hazard index scores), but Pfankuch channel stability scores were significantly higher for forested reaches compared with nonforested reaches. Furthermore, forested reaches had higher width to depth ratios, but flood-prone widths did not differ significantly, resulting in lower entrenchment ratios. Reduced channel width and cross-sectional area in nonforested reaches created an overall reduction of in-stream habitat, increased velocity, and increased potential for exceedance of channel capacity and floodplain access during high-flow events. Channel widening in response to riparian afforestation efforts has been observed in a variety of other locations globally and the results of this study suggest that widening with afforestation can still be anticipated in this region where stream gradients are low, hydrology is dominated by snowmelt, and forest cover is minimal.
Introduction
Riparian vegetation is used in bank stabilization, bank erosion control, restoring channel geomorphology, and habitat creation (Lyons et al. 2000). In comparison with forested stream reaches, nonforest reaches typically have lower channel widths, and this has been observed in multiple geographic locations including the Piedmont region of the United States (Zimmerman et al. 1967; Hession et al. 2003; Sweeney et al. 2004; McBride et al. 2008), the Midwestern USA (Trimble 1997; Zaimes and Schultz 2015), New Zealand (Davies-Colley 1997; Quinn et al. 1997), and the UK (Murgatroyd and Ternan 1983). Channel incision and reduction of overall stream habitat (i.e., benthic surface area) have generally been viewed as negative consequences of channel narrowing with deforestation in naturally forested regions (Sweeney et al. 2004). However, with riparian afforestation and the subsequent channel widening, unanticipated short-term changes in channel size and increases in sediment delivery downstream also may occur (Trimble 1997; McBride et al. 2008).
Most assessments of the impact of riparian vegetation on channel geomorphology have focused on regions outside the Canadian Prairies with higher topographic gradients, where native vegetation was primarily forest, and in regions where rainfall rather than snowmelt-driven runoff events are common (Lyons et al. 2000; Stewart et al. 2010). In other temperate regions, freeze–thaw cycles have been observed to increase potential for soil losses through bank failure or loosening of material, with highest rates frequently being observed under riparian forest (Zaimes and Schultz 2015). Although riparian soils on the prairies tend to remain frozen throughout the winter (Satchithanantham et al. 2017), bank thaw over the course of snowmelt and with inundation of flooded areas may be important processes defining bank erosion potential. Loosening of streambank soil during summer months following desiccation may also be an important factor influencing bank stability (Zaimes and Schultz 2015), and recent research into water use by riparian vegetation on the prairies suggests that water use is likely to be highest under riparian forests (Satchithanantham et al. 2017).
Although the physical drivers by which riparian vegetation and stream flow interact to shape channel geomorphology may be similar on the Canadian Prairies to those in other temperate agricultural regions (e.g., Zaimes and Schultz 2015), it has not yet been validated whether recommendations for riparian vegetation management that have been developed in other regions to increase bank stability or alter in-stream habitat are applicable for the prairie climate. In the current study we investigate the effect of vegetation type and health on channel geomorphology (cross-sectional characterization, bank erosion, and channel stability indicators) using paired forested and nonforest stream reaches located on the Canadian Prairies in southern Manitoba. Despite lower frequency of freeze–thaw cycles and greater dominance of annual hydrology by snowmelt on the prairies as compared with other regions, the results presented in this study indicate that wider channels are present with forested riparian areas and that similar processes define channel geomorphology to those observed in other regions.
Methods
Site selection
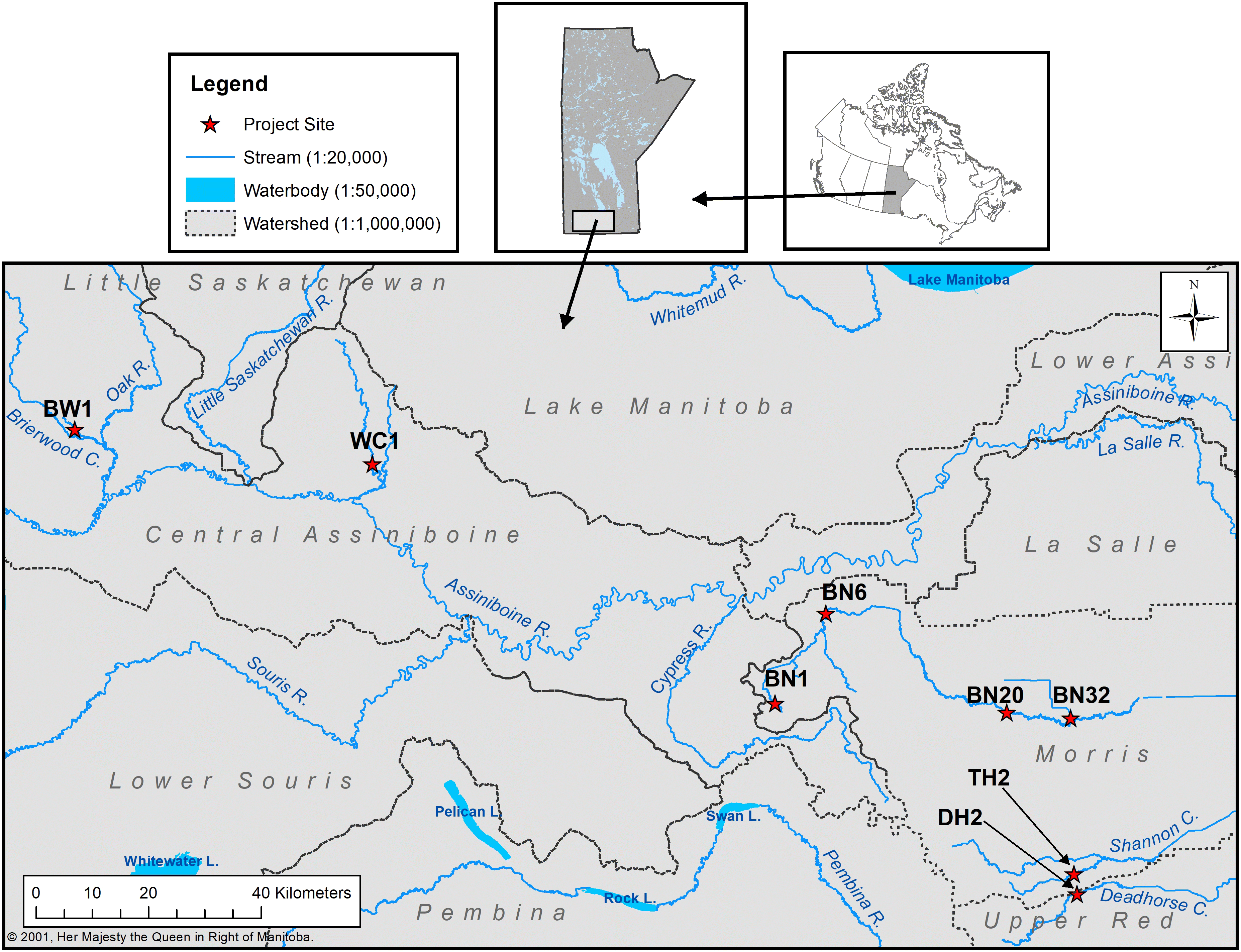
Using aerial photographs and site visits, the following criteria were utilized to select eight paired reaches in southern Manitoba (Fig. 1):
1.
The presence of one predominantly forested stream reach and one predominantly nonforested (grass and (or) pasture) stream reach at a site.
2.
The reach pairs are contiguous or nearly contiguous on the same stream.
3.
The length of each reach is 120–200 m long and representative of the general characteristics of the particular land use.
4.
No major tributaries enter within or between the reaches.
5.
No hydrological alterations such as culverts, berms, roads, or low level crossings are located within the reach.
The paired reach approach was chosen to ensure consistency of those characteristics other than vegetation type that influence channel form (soil, geology, topography, and climate).
Fig. 1.
Site descriptions
Sites were located in predominantly agricultural watersheds with annual cropland being the predominant land use (Table 1). Reach pairs were located on streams ranging from third order to fifth order according to the Strahler classification system (Table 1) (Manitoba Conservation and Water Stewardship 2018a). Figure 2 shows two paired reaches used in this study. The drainage area upstream of each paired site was delineated using a digital elevation model with either 1 or 15 m resolution, depending on data availability (Manitoba Conservation and Water Stewardship 2018b). Drainage areas ranged from approximately 42 to 917 km2 (Table 1). The forest reaches were upstream from the nonforest reaches at four sites and downstream at four sites. Regardless of whether the forest reaches were upstream of nonforest reaches or not, it was assumed that both reaches would have similar characteristics except vegetation. The average width of the stream corridors ranged from 24 to 336 m wide and the reach length ranged from 120 to 270 m. Three of the sites were located on a landscape with a gently rolling topography (average slope 5%–9%), and the other five sites were located on the low relief landscape of the Red River Valley (average slope 0%–2%). Soil type in the location of the drainage areas are categorized as black or grey Chernozems (Manitoba Conservation and Water Stewardship 2018c) (Table 2).
Table 1.
| Site ID | Stream | Stream ordera | Drainage area (km2) | Land cover in paired reaches (upstream/downstream) | Dominant land use in drainage areab |
|---|---|---|---|---|---|
| BN20 | Boyne River | 5 | 917 | Nonforest (grass)/forest | 63% annual cropland, 14% grassland/pasture, 13% trees |
| BN6 | Boyne River | 4 | 221 | Forest/nonforest (pasture) | 63% annual cropland, 15% grassland/pasture, 11% trees |
| DH2 | Deadhorse Creek | 4 | 128 | Nonforest (pasture)/forest | 71% annual cropland, 13% grassland/pasture, 8% trees |
| BW1 | Tributary to Brierwood Creek | 4 | 78 | Forest/nonforest (grass) | 67% annual cropland, 17% grassland/pasture, 4% trees |
| WC1 | West Branch Willow Creek | 3 | 80 | Nonforest (pasture)/forest (pasture) | 71% annual cropland, 14% grassland/pasture, 5% trees, 3% forages |
| TH2 | Thornhill Coulee | 3 | 68 | Nonforest (grass)/forest | 56% annual cropland, 22% grassland/pasture, 11% trees |
| BN32 | Tributary to Boyne River | 3 | 65 | Nonforest (grass)/forest | 75% annual cropland, 12% forages, 7% trees |
| BN1 | Tributary to Boyne River | 3 | 42 | Forest/nonforest (grass) | 73% annual cropland, 12% nonforestland/pasture, 6% trees |
a
Stream order was obtained from the Provincial Designated Drains, Manitoba Conservation.
b
Land use was determined using LandSat Imagery (BW1 site—2000 LandSat-7 Imagery; all other sites 2001 LandSat-5 Imagery).
Table 2.
| Site | EcoDistrict | Landscape | Drainage class and soil order | Annual precipitation (mm) | Climate |
|---|---|---|---|---|---|
| BN20 | MacGregor | Smooth, level to very gently sloping sandy glaciolacustrine plain with slopes that range from level to <2% | Imperfectly drained Gleyed Rego Black Chernozems | 500 | Cool, sub-humid boreal soil climate |
| BN6 | Stockton | Level to hummocky proglacial lacustrine plain | Well-drained Black Chernozemic soils | 490 | Cool, subhumid, boreal soil climate |
| BN32, DH2, TH2 | Winkler | Level to very gently sloping glaciolacustrine plain | Moderately well-drained to imperfectly drained Black Chernozems | 515 | Moderately cool, subhumid, boreal soil climate |
| DH2, TH2 | Manitou | Undulating to hummocky glacial till plain | Well-drained Black Chernozemic soils | 530 | Cool, subhumid, boreal soil climate |
| BW1, WC1 | Hamiota | Undulating to hummocky and kettled glacial till plain | Well-drained Black Chernozems | 460 | Moderately cold, subhumid, cryoboreal soil climate |
| WC1 | Shilo | Level to hummocky topography, with fine to coarse sands deposited by glacial meltwaters | Well-drained Black Chernozemic soil | 480 | Cool, subhumid, boreal soil climate |
| BN1, BN6, BN20 | Pembina | Strongly undulating and rolling to hummocky topography | Well-drained Dark Gray Chernozems, to a lesser extent, Gray Luvisols | 540 | Subhumid, moderately cold to cold, cryoboreal to boreal soil climate |
Fig. 2.

Field data collection
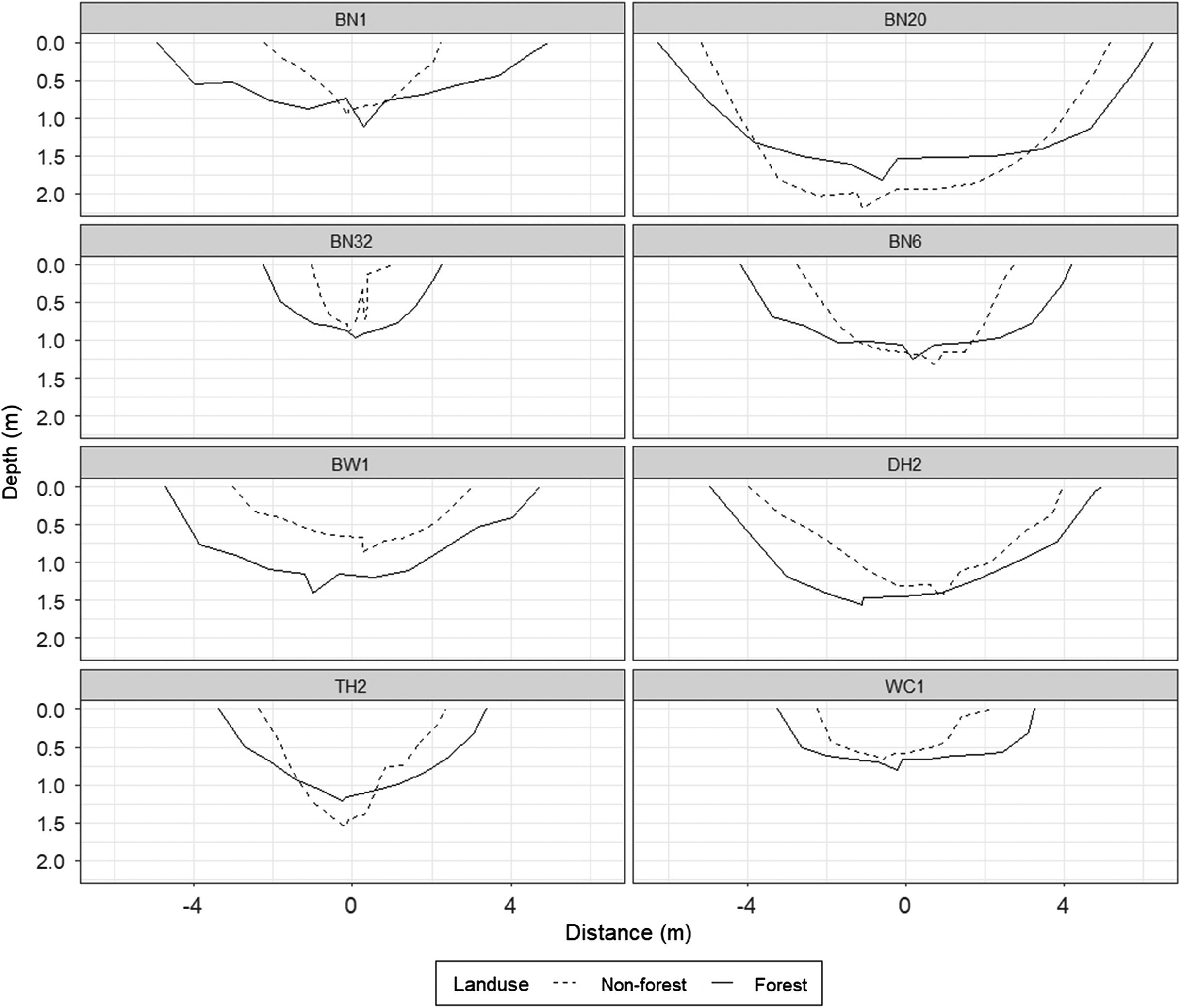
At each site, cross-sectional measurements of channel form were collected using methods described in Kline et al. (2004). Measurement transects were located at 20 m intervals along the reaches resulting in between 5 and 11 transects being measured per reach. Where possible, cross-sections were measured at locations where riffles are generally present.
At each cross-section, bankfull elevation was determined by visually studying bankfull indicators such as breaks in bank slope, tops of bars, and base of woody vegetation on stable banks. The bankfull elevation was used as a reference elevation since it can provide a more consistent measurement of channel dimensions that are comparable between sites. At each transect, bankfull depth measurements were taken across the channel at 10 equally spaced intervals and mean bankfull depth was calculated as the average of those 10 bankfull depths. This was accomplished by stringing a measuring tape horizontally across the channel at bankfull elevation and measuring the depth to channel bottom using a survey rod. The cross-sectional area of each transect was calculated by using the data from the cross-sectional survey measurements for each transect and an average cross-sectional area was calculated for each reach. Flood-prone width was measured at an elevation that corresponded to twice the maximum bankfull depth. Low bank height was determined as the height of the lowest bank, relative to bankfull maximum depth.
Other information collected at each site included large woody material, detritus, streambank soil texture, consistency of bank material, as well as streambank characteristics (bank height, root depth, root density, bank angle, and surface protection). The large woody debris (LWD) was estimated by counting the number of large pieces of wood that were found within the bankfull depth under each land use type. Only wood pieces that are at least 6 feet in length, 6 inches minimum diameter, and 12 inches diameter at the wider end were counted as LWD (Kline et al. 2004).
Indicators
From the field measurements, width to depth ratio (bankfull width to mean bankfull depth), entrenchment ratio (floodprone width to bankfull width), and incision ratio (maximum bankfull depth/low bank height) were calculated. Final values for each reach were determined by averaging the measurements at each of the cross-sections.
The Bank Erosion Hazard Index (BEHI) was developed by Rosgen (2001) to predict sediment contribution from stream banks in Montana and Colorado. Several physical characteristics of the bank were surveyed and used in estimating this multi-metric BEHI score, where stream banks are categorized into four ratings (low, moderate, high, and very high) based on erosion potential of the stream banks. Streambank variables collected for BEHI include bank height ratio (streambank height/maximum bankfull depth), rooting depth/bank height ratio, rooting density, percent surface area of bank protected by vegetation, and bank angle (Rathbun 2008).
The Pfankuch Channel Stability Scoring (PCSS) was developed by Pfankuch (1975) to evaluate overall stability of the channel and has been used in other parts of North America (Harmel et al. 1999). A numerical score was assigned to each of the 15 parameters that was evaluated in the PCSS, and the sum of the category ratings was transformed into a rating of reach condition by stream type (Rosgen 2001). Since this evaluation was developed for mountain streams, certain ratings were not relevant to the prairie streams investigated. In particular, the channel bottom category, four parameters (rock angularity, brightness, consolidation of particles, and bottom size distribution) were removed to make the PCSS more applicable. Similar to the modification made in Arizona (Jones 2015), a correction factor of 1.27 was used to convert the scores equivalent with the original Pfankuch scoring to categorize the channel stability. Sinuosity was calculated as the ratio of meandering channel length to the straight-line valley length using aerial imagery (Jones 2015).
The riparian area vegetation in each reach was characterized in greater detail using the Alberta Lotic Assessment (Fitch et al. 2009). The Alberta Lotic Assessment was simplified to include only the key components to accommodate the smaller size of the study sites and the modifications to the plant community classification (Fitch et al. 2009). The key components, identified as vegetation canopy complexity (Liu et al. 2010), presence and regeneration of tree and shrub species (Jones et al. 1994), and the percentage cover of invasive and disturbance plant species (Naiman et al. 2005), were surveyed and a streambank vegetation health score was estimated for each reach at each site. The methodology for compiling the data was taken from Fitch et al. (2009).
All statistical analysis was carried out with JMP (version 12, SAS Institute Inc., Cary, North Carolina, USA) with a probability of statistical significance tested at α = 0.05. The effect of vegetation type (forest vs. nonforest) on different geomorphic parameters was compared using paired t tests.
Results
The summary statistics (mean, ±standard deviation, and p-value of paired t test) of geomorphological characteristics for both forest and nonforest reaches are given in Table 3. The mean (±standard deviation) of LWD was significantly higher (p < 0.01) in forested reaches (7.5 ± 1.65 pieces per 100 m) compared with nonforest reaches (1.0 ± 0.65 pieces per 100 m). This pattern indicates that LWD largely originates from adjacent forested riparian areas. The smaller amount of LWD that was present in some nonforest reaches was likely transported from upstream locations during periods of high flow. Maximum bankfull depths and mean bankfull depths were not significantly different between forested and nonforested stream reaches. However, forest reaches were significantly wider (1.21–2.23 times wider, p < 0.01) and had 1.5 ± 0.4 times width to depth ratios (p < 0.05) compared with nonforest reaches (Table 3) and this pattern was observed regardless of upstream contributing drainage area (Fig. 3). The average cross-sectional area was also significantly higher (1.8 ± 0.6 times, p < 0.05) in forested reaches compared with nonforested reaches (Table 3, Fig. 4).
Table 3.
| Forest | Nonforest | Paired t test | ||||
|---|---|---|---|---|---|---|
| Parameter (units) | Mean | SD | Mean | SD | p-value | |
| LWD (pieces/100 m of stream) | 7.50 | 4.67 | 1.00 | 1.84 | <0.01 | |
| Detritus (%) | 39.19 | 16.98 | 24.69 | 17.12 | 0.05 | |
| Bankfull width (m) | 8.49 | 2.50 | 5.69 | 2.51 | <0.01 | |
| Mean bankfull depth (m) | 0.85 | 0.23 | 0.80 | 0.35 | 0.84 | |
| Width to depth ratio (m/m) | 10.53 | 3.09 | 7.65 | 2.60 | <0.05 | |
| Flood prone width (m) | 24.3 | 7.74 | 25.25 | 7.34 | 0.41 | |
| Entrenchment ratio (m/m) | 3.16 | 1.12 | 5.49 | 2.08 | <0.01 | |
| Incision ratio (m/m) | 1.60 | 0.43 | 1.53 | 0.27 | 0.59 | |
| Cross sectional area (m2) | 7.86 | 4.62 | 5.37 | 5.15 | <0.01 | |
| Sinuosity (m/m) | 1.5 | 0.35 | 1.25 | 0.30 | <0.05 | |
| Land form slope (unitless) | 5.77 | 1.95 | 4.17 | 1.58 | 0.05 | |
| Mass wasting (unitless) | 7.84 | 2.10 | 6.78 | 2.01 | 0.32 | |
| Debris jam potential (unitless) | 5.45 | 1.41 | 2.58 | 1.09 | <0.01 | |
| Vegetative bank protection (unitless) | 6.73 | 1.97 | 7.64 | 1.98 | 0.38 | |
| Channel capacity (unitless) | 1.14 | 0.20 | 1.14 | 0.31 | 0.96 | |
| Bank rock content (unitless) | 8.00 | 0.00 | 7.63 | 0.74 | 0.20 | |
| Obstructions to flow (unitless) | 4.68 | 1.00 | 3.53 | 1.08 | <0.05 | |
| Cutting (unitless) | 10.31 | 2.48 | 10.00 | 3.17 | 0.72 | |
| Deposition (unitless) | 9.03 | 1.61 | 7.92 | 3.07 | 0.24 | |
| Scouring and deposition (unitless) | 14.15 | 3.48 | 14.25 | 2.49 | 0.92 | |
| Aquatic vegetation (unitless) | 3.03 | 1.00 | 2.65 | 0.77 | 0.35 | |
| PCSS (unitless) | 96.56 | 13.46 | 86.61 | 13.29 | <0.01 | |
| Root depth (m) | 0.41 | 0.11 | 0.28 | 0.08 | <0.05 | |
| Bank height (m) | 2.12 | 1.10 | 1.90 | 1.04 | 0.10 | |
| Root density (%) | 57.47 | 15.10 | 48.70 | 17.53 | 0.08 | |
| Bank surface protection (%) | 57.68 | 13.68 | 59.06 | 16.95 | 0.85 | |
| Lower bank angle (°) | 71.48 | 10.10 | 75.21 | 9.79 | 0.29 | |
| Maximum bankfull height (m) | 1.27 | 0.32 | 1.24 | 0.50 | 0.80 | |
| Bank height/max bankfull depth (m/m) | 1.60 | 0.41 | 1.51 | 0.29 | 0.48 | |
| Root depth/bank height (m/m) | 0.23 | 0.12 | 0.18 | 0.10 | 0.20 | |
| Weighted root density (unitless) | 12.87 | 5.53 | 8.90 | 6.00 | <0.05 | |
| Bank Erosion Hazard Index (unitless) | 29.37 | 3.82 | 30.67 | 2.98 | 0.39 | |
| Vegetation assessment score (unitless) | 18.25 | 3.11 | 11.25 | 3.96 | <0.01 | |
Fig. 3.
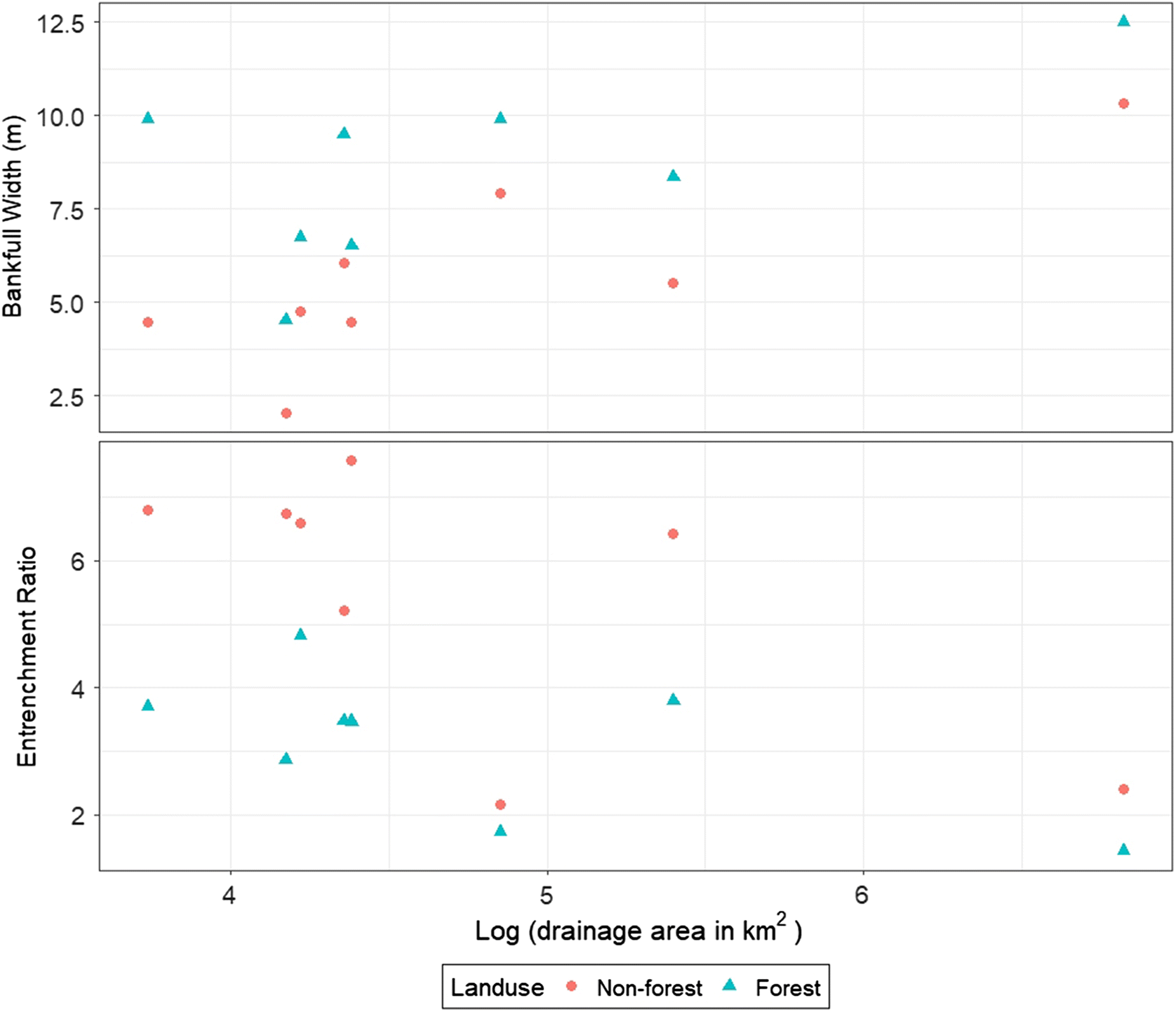
Fig. 4.
In the prairie study region the vegetative bank protection rating (measured as a part of the PCSS and shown in Table 3) was slightly higher in nonforest reaches (7.64 ± 0.70) compared with forested reaches (6.73 ± 0.70), although that difference was not significant and the overall stream bank vegetation health score measured using the modified Alberta Lotic assessment was significantly higher in forest reaches compared with nonforest reaches (p < 0.01). Only two sites (BN6 and TH2) actually exhibited poorer vegetative bank protection in nonforest reaches as compared with forest reaches and BN6 was the only site showing impact from livestock grazing. BEHI was marginally lower in forest reaches; however, differences between forest and nonforest reaches were not statistically significant.
Discussion
The trend in LWD observed in Manitoba stream reaches was similar to those that have been observed in naturally forested regions, but it was lower in magnitude. For example, in Vermont, USA, nonforested reaches were almost without LWD whereas forested reaches had an average of 14.2 pieces per 100 m of reach length (McBride et al. 2008) and Quinn et al. (1997) reported that compared with pasture, pine forest had 17 times higher LWD volume. The number of LWD found in smaller streams is a function of age of the riparian forest where transport of LWD is minimal (McBride et al. 2008). Although trees present in the current study were all well established, the age of these stands was not defined. The reason for the low magnitude of LWD in the current study could be due to differences in age and species of the trees. Reaches where trees have only recently been established are likely to have fewer pieces of LWD, but rates of accumulation over time remain to be defined for tree species typical of the prairies.
Stream width and bank stability
Streams with cohesive bank material (e.g., clay) tend to be deeper while channels with noncohesive bank material (e.g., silt and sand) tend to be wider and shallower (Baillie and Davies 2002); however, the paired reach design of the current study allowed for some control over this effect and for detection of vegetation specific differences in channel form. The width to depth ratio can be used as an indicator to gauge the degree of control on stream channel capacity by vegetation rather than using flow-resistance guides. According to Masterman and Thorne (1992), stream reaches with width to depth ratio of <16 have greater influence by vegetation on channel capacity than reaches with higher ratios. In this study, with the exception of one forested reach, all reaches had width to depth ratios <16, indicating significant influence of vegetation on flow capacity of the channels.
Wider bankfull widths (Zimmerman et al. 1967; Quinn et al. 1997; Sweeney et al. 2004) and higher cross-sectional areas (Trimble 1997; Baillie and Davies 2002) in reaches have been observed for a variety of regions outside the Canadian Prairies where native riparian vegetation is primarily forest. Within a region, width and cross-sectional area tend to scale with watershed area (Bieger et al. 2015) and this trend was observed for the current study, where, in all cases, width was higher for forested reaches (Fig. 3). In the northern Vermont region Zimmerman et al. (1967) observed no effect of vegetation on channel form once catchment size exceeded 13 km2 (Zimmerman et al. 1967). Anderson et al. (2004) noted that in larger watersheds (>10 to 100 km2) nonforested reaches were wider than forested reaches, though this trend reverses for smaller watersheds where the equivalent threshold in terms of channel width is 20 m. It should be noted that in the current study, all the reaches have average widths less than this threshold with a maximum width of 12.5 m. Furthermore, differences between reaches with forest and nonforest vegetation were also observed for larger watershed areas; however, the largest differences tended to occur in smaller watersheds (Fig. 3). Lower annual water yield on the Canadian Prairies versus Vermont will typically result in lower rates of flow for a given watershed area and may partially explain why these effects of vegetation on channel form are also observed in larger watersheds in the current study.
The lack of statistical difference in erosion potential (measured by BEHI) indicates that the potential sediment transport for both vegetation types could be quite similar, regardless of the differences in LWD and channel width, once the channel is in equilibrium. However, it is still possible to have higher sediment delivery from forest reaches during the channel widening process.
Potential implications of changes in channel form for stream hydrology and sediment dynamics
Patterns observed across a variety of other study regions and in the current study area suggest stream widening can be anticipated with reforestation of riparian areas that are currently dominated by grass vegetation, and narrowing is likely to occur following deforestation. This pattern of reduced channel width in reaches dominated by grasses has been attributed to higher cohesiveness of bank material where the root system of grasses is present at a high density with greater bank stabilization and reduced erosion (Murgatroyd and Ternan 1983; Davies-Colley 1997). Reaches with grass vegetation may have a higher amount of sediment stored in their banks compared to similar reaches with trees (Davies-Colley 1997; Quinn et al. 1997). Where reforestation occurs, nonforest reaches have the potential for high sediment yield during the processes of channel adjustment, widening, and slumping of bank material and can lead to sedimentation of channel bed and increased turbidity of the water (Davies-Colley 1997; McBride et al. 2008).
A significantly lower entrenchment ratio in forest reaches (Fig. 4, p < 0.01) indicates that channels in forested areas were less likely to have access to the floodplain. Due primarily to lower channel widths, nonforested reaches have smaller cross-sectional areas and it can be anticipated that these reaches may tend to spill overbank more frequently in comparison to forested reaches. Given the narrower cross-sectional areas of nonforested reaches it is possible that under high-flow conditions higher stream velocities could result in the supply of more sediment from channel incision in nonforested reaches than forested sites. Most of the reaches included in the current study are slightly entrenched (entrenchment ratio > 2.2) and have a low width/depth ratio that is <12. In North Dakota (Meehan et al. 2016) and in Ohio (Ward et al. 2008), where similar geophysical conditions to Manitoba exist, streams had a similar channel form.
Assuming similarities between the Manitoba streams studied here and streams in other regions with similar channel form, some insight can be gained into the likelihood for erosion, sediment transport, and deposition. Similar stream types tend to have very high sensitivity to disturbance, good recovery potential, low sediment supply, moderate stream bank erosion supply, and very high vegetation controlling influence (Brooks et al. 2012). Extending the study to other areas of Manitoba and by increasing the sample size, the confidence of this prediction can be increased.
Similar studies could be carried out to determine if a reach is degraded and provide information to support whether restoration is warranted or not. For example, planting trees during restoration may contribute to the widening of the channel and higher sediment yield than prerestoration levels. So the vegetation effects on geomorphology should be viewed separately from the geological effects. The results of the study provide insights into what kind of change/adjustments in channel form could be expected if riparian reforestation is carried out. Also, with additional work, geomorphic metrics presented in this study could be used to predict sediment transport or to develop new relationships specific for the prairie region. If stream bank erosion is measured, the results can be used in conjunction to predict sediment delivery.
Research examining the implications of reforestation of riparian zones over time suggests that after an initial period of channel adjustment a new equilibrium is achieved through aggradation or development of an inset floodplain (McBride et al. 2008). Prairie sites included in the current study had relatively well-established tree and grass stands, but whether these represent a relatively stable channel state is not a question that could be addressed without multi-year data. However, the tendency toward wider channel, lower potential for floodplain inundation, and increases in LWD accumulation under established stands of trees that has been observed in other regions is present in the Canadian Prairie region. Given the similarities in trends between regions, it is possible to adapt riparian management recommendations for control of channel form from other regions.
Conclusion
Mean bankfull width, LWD, detritus, width to depth ratio, cross-sectional area, sinuosity, land form slope, debris jam potential, obstruction to flow, root depth, PCSS, weighted root density, and streambank vegetation health were significantly higher in forest reaches compared with nonforest reaches. These patterns observed in the current study suggest that if riparian bank stabilization and restoration efforts on the Canadian Prairies result in a shift from grass to tree vegetation, the impact on channel form and sediment dynamics may be very similar to that observed in other regions. In Prairie streams where bank material is noncohesive, with a shift from grass to trees, sediment yield may increase initially while the channel increases in width. However, on stream channels where riparian trees are well established, the flow capacity of stream channels tends to be greater, the area of in-stream habitat increased, travel times longer in comparison with nonforest reaches, and the potential for exceedance of channel capacity and floodplain access during high-flow events is reduced. Further research is required to better define how changes associated with differing riparian vegetation might be anticipated to occur through time. Furthermore, the stream reaches included in the current study were primarily nongrazed to avoid the confounding influence of differing intensities of disturbance. Additional research is required to define how the influence of vegetation type on channel form and erodibility is likely to be impacted by grazing disturbance in the region. Streambank vegetation is naturally mixed on the Canadian Prairies and was historically influenced by flooding, fire, and grazing. This study indicates that forest and nonforest vegetation types on the Canadian Prairies are each likely to offer benefits and trade-offs at differing timescales for channel stability, sediment delivery, in-stream habitat, and channel capacity.
Acknowledgements
This research was supported by funding under Agriculture and Agri-Food Canada’s Growing Forward 2 Program. The authors would like to acknowledge the technical support provided by AAFC employees Blair English, Mike Chubey, and Greg Gatsky and to thank landowners who cooperated with the field crew to allow for access to field sites. Feedback received from two anonymous reviewers significantly improved this manuscript.
References
Anderson RJ, Bledsoe BP, and Hession WC. 2004. Width of streams and rivers in response to vegetation, bank material, and other factors. Journal of the American Water Resources Association, 40(5): 1159–1172.
Baillie BR, and Davies TR. 2002. Effects of land use on the channel morphology of streams in the Moutere Gravels, Nelson, New Zealand. Journal of Hydrology, 41(1): 19–45.
Bieger K, Rathjens H, Allen PM, and Arnold JG. 2015. Development and evaluation of bankfull hydraulic geometry relationships for the physiographic regions of the United States. Journal of the American Water Resources Association, 51(3): 842–858.
Brooks KN, Ffolliott PF, and Magner JA. 2012. Fluvial processes and implications for stream management. Chapter 10. In Hydrology and the management of watersheds. Edited by KN Brooks, PF Ffolliott, and JA Magner. John Wiley & Sons, Inc., New York, New York. pp. 267–295.
Davies-Colley RJ. 1997. Stream channels are narrower in pasture than in forest. New Zealand Journal of Marine and Freshwater Research, 31(5): 599–608.
Fitch L, Adams BW, and Hale G. 2009. Riparian health assessment for streams and small rivers—field workbook. Cows and Fish Program, Lethbridge, Alberta. 94 p.
Harmel RD, Haan CT, and Dutnell RC. 1999. Evaluation of Rosgen’s streambank erosion potential assessment in northeast Oklahoma. Journal of the American Water Resources Association, 35(1): 113–121.
Hession WC, Pizzuto JE, Johnson TE, and Horwitz RJ. 2003. Influence of bank vegetation on channel morphology in rural and urban watersheds. Geology, 31(2): 147–150.
Jones CG, Lawton JH, and Shachak M. 1994. Organisms as ecosystem engineers. In Ecosystem management. Edited by FB Samson and FL Knopf. Springer, New York, New York. pp. 130–147.
Jones JD. 2015. Standard operating procedures for surface water quality sampling. Surface Water Section, Arizona Department of Environmental Quality (ADEQ), Phoenix, Arizona.
Kline M, Alexander C, Pomeroy S, Cahoon B, and Becker L. 2004. Vermont stream geomorphic assessment: phase 2 handbook. Vermont Agency of Natural Resources, Waterbury, Vermont.
Liu D, Diplas P, Hodges CC, and Fairbanks JD. 2010. Hydrodynamics of flow through double layer rigid vegetation. Geomorphology, 116(3–4): 286–296.
Lyons J, Thimble SW, and Paine LK. 2000. Grass versus trees: managing riparian areas to benefit streams of central North America. Journal of the American Water Resources Association, 36(4): 919–930.
Manitoba Conservation and Water Stewardship. 2018a. Designated drain watercourses [online]: Available from mli2.gov.mb.ca/water_resources/des_drain_index.html.
Manitoba Conservation and Water Stewardship. 2018b. Digital elevation models (DEMs) [online]: Available from mli2.gov.mb.ca/dems/index.html.
Manitoba Conservation and Water Stewardship. 2018c. Soil mapping by municipality [online]: Available from mli2.gov.mb.ca/soils/index.html.
Masterman R, and Thorne CR. 1992. Predicting influence of bank vegetation on channel capacity. Journal of Hydraulic Engineering, 118(7): 1052–1058.
McBride M, Hession WC, and Rizzo DM. 2008. Riparian reforestation and channel change: a case study of two small tributaries to Sleepers River, northeastern Vermont, USA. Geomorphology, 102(3–4): 445–459.
Meehan MA, Sedivec KK, Hecker GA, and Printz JL. 2016. Riparian complex ecological sites of North Dakota: a pictorial guide of riparian complex ecological sites common in North Dakota [online]: Available from ag.ndsu.edu/pubs/ansci/range/r1791.pdf.
Murgatroyd AL, and Ternan JL. 1983. The impact of afforestation on stream bank erosion and channel form. Earth Surface Processes and Landforms, 8: 357–369.
Naiman RJ, Décamps H, and McClain ME. 2005. Riparia: ecology, conservation, and management of stream side communities. Elsevier B.V., Burlington, Massachusetts.
Pfankuch DJ. 1975. Stream reach inventory and channel stability evaluation. U.S. Department of Agriculture Forest Service, Missoula, Montana. 26 p.
Quinn JM, Cooper AB, Davies-Colley RJ, Rutherford JC, and Williamson RB. 1997. Land use effects on habitat, water quality, periphyton, and benthic invertebrates in Waikato, New Zealand, hill-country streams. New Zealand Journal of Marine and Freshwater Research, 31(5): 579–597.
Rathbun J. 2008. Standard operating procedure: assessing bank erosion potential using Rosgen’s Bank Erosion Hazard Index. Michigan Department of Environmental Quality, Water Bureau, Nonpoint Source Unit, Lansing, Michigan.
Rosgen DL. 2001. A practical method of computing streambank erosion rate. In Proceedings of the Seventh Federal Interagency Sedimentation Conference, Reno, Nevada, 25–29 March 2001. pp. 9–17. Available from https://semspub.epa.gov/work/01/554370.pdf.
Satchithanantham S, Wilson HF, Glenn AJ. 2017. Contrasting patterns of groundwater evapotranspiration in grass and tree dominated riparian zones of a temperate agricultural catchment. Journal of hydrology, 549: 654–666.
Smith RE, Veldhuis H, Mills GF, Eilers RG, Fraser WR, and Lelyk GW. 1998. Terrestrial ecozones, ecoregions and ecodistricts, an ecological stratification of Manitoba’s natural landscapes. Technical Bulletin 98-9E. Land Resource Unit, Brandon Research Centre, Research Branch, Agriculture and Agri-Food Canada, Winnipeg, Manitoba. 324 p.
Stewart A, Reedyk S, Franz B, Fomradas K, Hilliard C, and Hall S. 2010. Field manual on buffer design for the Canadian Prairies. Agri-Environmental Services Branch, Agriculture and Agri-Food Canada, Ottawa, Ontario.
Sweeney BW, Bott TL, Jackson JK, Kaplan LA, Newbold JD, Standley LJ, et al. 2004. Riparian deforestation, stream narrowing, and loss of stream ecosystem services. Proceedings of the National Academy of Sciences of the USA, 101(39): 14132–14137.
Trimble SW. 1997. Stream channel erosion and change resulting from riparian forests. Geology, 25(5): 467–469.
Ward A, D’Ambrosio JL, and Mecklenburg D. 2008. Stream classification. Agriculture and Natural Resources Fact Sheet AEX-445-01. The Ohio State University Extension, Columbus, Ohio. pp. 1–8.
Zaimes GN, and Schultz RC. 2015. Riparian land-use impacts on bank erosion and deposition of an incised stream in north-central Iowa, USA. Catena, 125: 61–73.
Zimmerman RC, Goodlett JC, and Comer GH. 1967. The influence of vegetation on channel form of small streams. In Symposium on River Morphology. International Association of Hydrological Sciences, Gentbrugge, Belgium. General Assembly of Bern, Switzerland (25 September–7 October 1967). pp. 255–275 [online]: Available from http://www.uvm.edu/~pbierman/classes/gradsem/2005/papers/zimm1967.pdf.
Information & Authors
Information
Published In

FACETS
Volume 4 • Number 1 • June 2019
Pages: 336 - 349
Editor: Jeremy Kerr
History
Received: 11 September 2018
Accepted: 1 May 2019
Version of record online: 31 July 2019
Copyright
© 2019 Satchithanantham et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Key Words
Sections
Subjects
Authors
Author Contributions
SS and HFW conceived and designed the study.
SS, PM, and MD performed the experiments/collected the data.
SS, HFW, PM, and MD analyzed and interpreted the data.
HFW contributed resources.
SS, HFW, SL, and AJK drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Sanjayan Satchithanantham, Henry F. Wilson, Patsy Michiels, Melanie Dubois, Sheng Li, and Alexander J. Koiter. 2019. Channel geomorphology differences between stream reaches with grass- or tree-dominated riparian vegetation in southern Manitoba. FACETS.
4(1): 336-349. https://doi.org/10.1139/facets-2018-0036
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Putting down roots: Afforestation and bank cohesion of Icelandic Rivers