Recommended best practices for plastic and litter ingestion studies in marine birds: Collection, processing, and reporting
Abstract
Marine plastic pollution is an environmental contaminant of significant concern. There is a lack of consistency in sample collection and processing that continues to impede meta-analyses and large-scale comparisons across time and space. This is true for most taxa, including seabirds, which are the most studied megafauna group with regards to plastic ingestion research. Consequently, it is difficult to evaluate the impacts and extent of plastic contamination in seabirds fully and accurately, and to make inferences about species for which we have little or no data. We provide a synthesized set of recommendations specific for seabirds and plastic ingestion studies that include best practices in relation to sample collection, processing, and reporting, as well as highlighting some “cross-cutting” methods. We include guidance for how carcasses, regurgitations, and pellets should be handled and treated to prevent cross-contamination, and a discussion of what size class of microplastics can be assessed in each sample type. Although we focus on marine bird samples, we also include standardized techniques to remove sediment and biological material that are generalizable to other taxa. Lastly, metrics and data presentation of ingested plastics are briefly reviewed in the context of seabird studies.
Introduction
Seabirds are particularly vulnerable to ingesting marine plastic pollution. Most seabird species feed at or near the ocean surface, the same zone in which marine plastics occur at a high density, along eddies and oceanic convergence zones, mistaking the plastics for prey items (Cadée 2002; Janinhoff et al. 2010; Law et al. 2010; Song et al. 2014; Reisser et al. 2015). Ingestion of plastic was first reported in seabirds in the 1960s (Threlfall 1968; Kenyon and Kridler 1969), and there is a now a relatively comprehensive and widespread body of literature for seabirds compared with other marine megafauna groups (Provencher et al. 2017). Most of this work has focused on recording the type and number of items ingested by seabirds (i.e., frequency of occurrence; Provencher et al. 2017), with plastics reported in >200 species of seabirds (Kühn et al. 2015).
Currently the only species (of any biota) where a published, standardized protocol has been adopted to assess plastic ingestion is the seabird the northern fulmar (Fulmarus glacialis (Linnaeus, 1761)) (van Franeker et al. 2011; van Franeker and Law 2015; Provencher et al. 2017). These methods have been applied successfully in the broader North Atlantic region as well as in the North Pacific Ocean and have enabled some of the few, ocean-scale comparisons of marine plastic pollution over time and space (van Franeker et al. 2011; Provencher et al. 2017; Avery-Gomm et al. 2018). Although these methods of quantifying plastic pollution are often used elsewhere, most studies fail to report plastic ingestion results in a comprehensive way (O’Hanlon et al. 2017; Provencher et al. 2017).
Standardized reports of plastic ingestion (or lack of observed plastic ingestion) are important because they: (1) highlight the magnitude of the plastic pollution issue; (2) contribute to meta-analyses that report temporal, spatial, and taxonomic trends in plastic ingestion; and (3) could help elucidate the population level effects of plastic ingestion via correlative studies of load and demographic rates. Note that the absence of plastics in species is an important component of these standardized approaches (Provencher et al. 2017). Additionally, among species that are suitable as biological indicators of marine plastic pollution, reporting can provide a cost-efficient way to monitor the progress of waste management or plastic source reduction strategies that aim to reduce marine plastic pollution.
Unfortunately, a lack of standardized methods for sample collection along with comparable processing and reporting can undermine broad-scale assessments of the extent and impact of marine plastic debris on marine biota. The importance of lab processing methods was recently illustrated when two studies documenting plastic ingestion in birds from the same stranding event could not be compared because they followed different methods (Fife et al. 2015; Avery-Gomm et al. 2016). Additionally, some studies have not processed birds and presented data in the same way, such as reporting differences between sexes or among age classes (Provencher et al. 2014; Roman et al. 2016). Given that data on plastic and debris ingestion by seabirds form some of the most comprehensive datasets on marine pollution in terms of spatial, temporal, and phylogenetic coverage, there is a need to quickly standardize sample collection, processing, plastic quantification and reporting so as to improve the integrity of plastic ingestion research, and will facilitate statistically and scientifically credible large-scale comparisons in the future.
As plastic reduction policies are developed and implemented, directly comparable data are needed to test the efficacy of these policies at reducing plastic pollution in biota. Data from northern fulmars in the North Sea were used to test policy targets for plastic reduction, and have since helped policy makers and researchers evaluate if the policy goals were realistic and achievable (van Franeker et al. 2011; OSPAR 2015, 2017; Provencher et al. 2017). With a growing recognition of the importance of evidence-driven policy, consistent and repeatable monitoring methods and approaches are needed to evaluate reductions in the abundance of plastics in the world’s oceans (Borrelle et al. 2017).
We provide a synthesized set of recommendations specific for seabirds and plastic ingestion studies that include best practices in relation to sample collection, processing methods, and reporting. We include standardized methods for assessing plastic ingestion by seabirds using necropsy, regurgitations, and pellets. Although each method has advantages and disadvantages (Table 1), each is appropriate in some scenarios. Adopting this approach will move the global plastic research community towards addressing some of the key questions about how or why species are affected by plastics, and other global research objectives that extend beyond the jurisdiction of a single species or study.
Table 1.
| Pros | Cons | |
|---|---|---|
| Necropsy—carcass examination (dissection) | - Can determine entire plastic burden - Tissues can also be easily taken for plastics-associated contaminants analysis - Sex and age, health status, and cause of death can be determined - In case of beach-cast birds: non-invasive | - Does not always allow for consistent repeated sampling - Is usually opportunistic without options for preplanned sampling in relation to season, sex or age - In the case of hunting specifically for research samples this technique is invasive |
| Regurgitations | - Can be repeated on the same individuals to compare plastic load between seasons and years - In comparison to hunting birds for research, regurgitations are relatively less invasive | - Not all species can be made to regurgitate - Does not guarantee a complete sample - Low but real possibility of injury occurring to the bird - The individual may lose a meal that was costly to acquire |
| Pellet collection | - Easily collected - Non-invasive collections - Can be repeated between seasons and across years - Can be regularly sampled - Can be implemented easily and at low cost | - Cannot be used to assess microplastics - Depending on the species, may be difficult to attribute pellets to individuals - Does not guarantee a complete sample - Typically only useful during the breeding season when birds are occupying nesting sites on land |
Methods
We define seabirds to include gulls, terns, skimmers, skuas, auks, and phalaropes (Charadriiformes), tropicbirds (Phaethontiformes), penguins (Sphenisciformes), petrels, shearwaters, and albatrosses (Procellariiformes), cormorants, frigatebirds, boobies, and gannets (Suliformes), and pelicans (Pelecaniformes) consistent with the definition proposed by Gaston (2004), with some updated classifications (Boesman et al. 2017). Although we focus on marine birds, these methods are applicable to all birds, particularly other aquatic species such as loons (Gaviiformes) and waterfowl (Anseriformes), in which plastic has been recorded (Avery-Gomm et al. 2013; Provencher et al. 2014; English et al. 2015; Holland et al. 2016; Gil-Delgado et al. 2017), and much is transferable to studies of ingested plastics in other vertebrates (e.g., marine mammals, turtles, and fish).
Sample collection in the field
Collection of intact carcasses
Dissection of carcasses is the most commonly used technique for assessing ingested plastics in seabirds (Provencher et al. 2017), and has several advantages over the other methods described here, including the ability to determine age/sex of individuals, potential internal pathology related to plastic burdens, and the most accurate measurement of plastic particles (van Franeker et al. 2011; Acampora et al. 2014; Jiménez et al. 2015; Box 1; Table 1). Several different measures of body condition can be examined, which is useful when exploring the potential implications of plastic ingestion on the health of biota including pectoral muscle size, body mass, and subcutaneous fat (Fry et al. 1987; Harper and Fowler 1987; van Franeker et al. 2011; Donnelly-Greenan et al. 2014). Carcasses can also readily permit sampling of internal tissues for subsequent pathological or chemical analyses (Lavers et al. 2014; Fife et al. 2015; Herzke et al. 2016), and in some jurisdictions permits and licenses are easier to obtain for salvaging carcasses than working with live birds.
Box 1. Key metrics to measure and report in seabird plastic ingestion studies.
• Species
• Age
• Sex
• Body condition metrics: pectoral muscle size, body mass (when the bird is dry and clean), and subcutaneous fat
• Cause of mortality (with necropsy, and where possible)
• Report zeros
Carcasses can come from a variety of sources, including beached birds, fisheries bycatch, ship strikes, rehabilitation centres, collaboration with hunters, as well as collection for scientific research (Provencher et al. 2017). The disadvantage of carcass collection is that it is generally opportunistic. Carcasses can often only be collected at irregular time intervals, which may be limited to species that are most likely to be beach-cast or hunted (Avery-Gomm et al. 2012; Provencher et al. 2014; Seif et al. 2018).
Although all carcasses should be treated in the same way when examined for ingested plastics, it is important to consider the source of mortality as it can influence the levels of plastics reported. The cause for mortality may or may not represent a true sample of the population and, thus, may affect how the results are interpreted. Plastic ingestion data may be biased in beached birds or those that died of unknown causes, as plastic may have contributed to the death of the animal (Auman et al. 1998) or have been a consequence of its poor condition. Although there were no significant differences in plastic burdens between beached northern fulmars and those that died in good condition (as a consequence of e.g., drowning in nets or by collision with ship cables, lights and antenna cables, etc.; van Franeker and Meijboom 2002), Short-tailed shearwaters (Ardenna tenuirostris (Temminck, 1835)) differed in their plastic burden depending on how they died (Rodríguez et al. 2018).
Regurgitations from birds
Some species can be made to regurgitate items in the field (Fig. 1), and 17% of published papers reported data on ingested plastics in seabirds using regurgitations, water-offloading (lavage or flushing), or emetics (Provencher et al. 2017). An advantage of regurgitations is that they provide a sample of items consumed very recently. Regurgitations also allow targeted sampling of species that are not commonly found as carcasses (i.e., non-hunted species and those not affected by fisheries bycatch or rarely found beached), and can provide particularly valuable insights into individual behaviour or preferences. Targeted sampling allows dedicated study designs with pre-determined sample sizes, focusing on particular age classes, and repeated sampling of individuals.
Fig. 1.
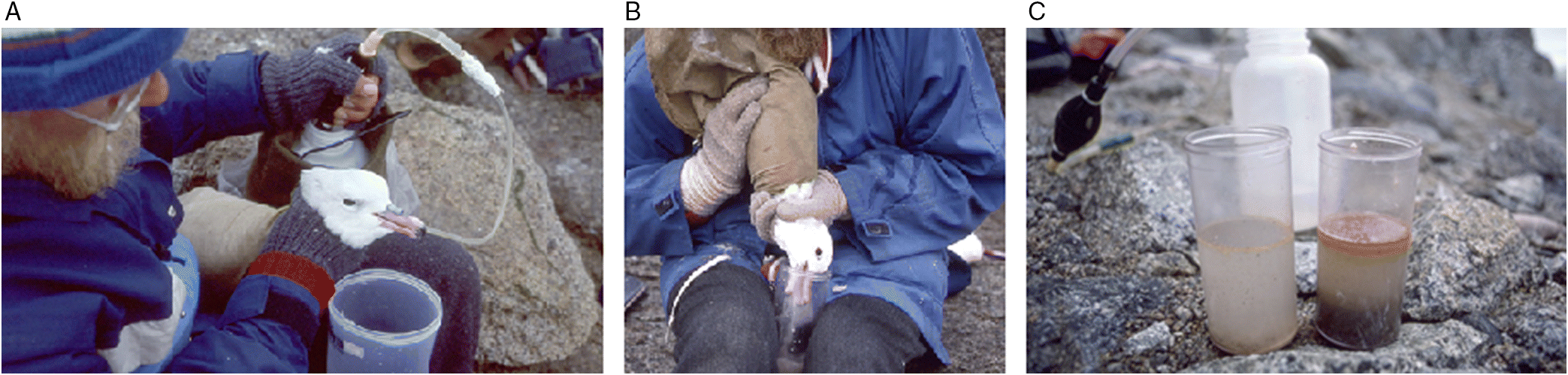
The main challenge with this suite of sampling techniques is acquiring a complete sample. Fijn et al. (2012) found considerable differences between diet data obtained from birds that had first regurgitated spontaneously, and were subsequently lavaged. Lavage likely returns the complete proventricular contents (Fig. 2), but it is less clear whether the entire contents of the gizzard are flushed out. In Procellariiformes, gizzard contents may not be regurgitated (Furness 1985; Ryan and Jackson 1986), whereas gizzard contents were regurgitated completely when using emetics (Bond and Lavers 2013). Between 6% and 22% of plastic items remained inside shearwater fledglings after lavage (Hutton et al. 2008; Lavers et al. 2014), particularly in individuals with large amounts of plastic, and therefore mass and frequency data collected using this technique should be considered underestimates in some species (Bond and Lavers 2013). Lavage and emetics may be ineffective in some groups of seabirds, such as those that do not regurgitate food to offspring (e.g., auks, but see Wilson et al. 2004). The completeness of the sample likely depends on the species’ stomach morphology, as different species have differently shaped gastrointestinal tracts (GITs; Fig. 2), and the methods applied.
Fig. 2.

The impacts of stomach lavage on seabirds are species specific. In some penguins, there were no negative effects of stomach lavage on adult survival, chick growth, or breeding success, and the procedure did not alter the foraging behaviour of adults (Robertson et al. 1980; Goldsworthy et al. 2016). However, in diving petrels there was 2.3% mortality (5 out of 220 handled birds; Jahncke et al. 1999) following lavage, and in Leach’s storm-petrels (Oceanodroma leucorhoa (Vieillot, 1818)), 8% mortality occurred when emetics were used (Bond and Lavers 2013). In 323 stomach flushings of four species of Antarctic fulmarine petrel and one southern fulmar (Fulmarus glacialoides (Smith, 1840)), 0.3% died because a large prey item became stuck in the throat and caused shock. In breeding pairs with chicks, sampling was restricted to one parent during the season, which had no detectable effect on adult foraging behaviour or chick growth until fledging (van Franeker et al. 2001). Although lavage and emetics both require training and have an associated risk of harm to the animal if not done correctly (e.g., aspiration of fluid into the lungs), both can be useful methods in specific cases (Gaunt and Oring 1997; Bond and Lavers 2013).
Many birds regurgitate immediately upon being captured and, therefore, can be sampled using this technique relatively easily (Bond et al. 2010). However, not all individuals will regurgitate, or they will do so only partially, which makes distinguishing individuals with a true absence of plastic from those who contain plastic but did not regurgitate it difficult. For these species lavage or administering emetics may be preferable. For example, spontaneous regurgitations by gull chicks were incomplete and, therefore, underestimated the amount of ingested plastic to an unknown extent (Hunt 1972). Plastic burden may also be underestimated in spontaneous regurgitations if samples are collected from the ground, or plastic sheets placed below mist-nets, where small pieces of plastic may not be detected. For example, in a sampling of 973 Leach’s storm-petrels using spontaneous regurgitation between 1987 and 1988 and 2002 and 2006 from a single site in Newfoundland, 5%–6% contained plastic (Provencher et al. 2014). However, the same population sampled in 2012 using emetics (n = 63) was found to have a 48% frequency of occurrence of plastics, with all pieces <17 mg (Bond and Lavers 2013).
Collection of pellets
Several species regularly regurgitate pellets composed of indigestible items. These pellets or boluses can be found around nesting sites (Fig. 3), and 10% of plastics studies between 1965 and 2016 used this technique (Provencher et al. 2017). Examining pellets is a useful, non-destructive sampling technique (Lindborg et al. 2012; Hammer et al. 2016), and has been used most frequently for scavenging species, such as gulls and skuas. It can also be suitable for other seabirds such as cormorants, shags, and terns that produce pellets (Barrett et al. 2007; Álvarez et al. 2018). However, some species known to have the highest level of plastic accumulation (e.g., Procellariiformes) do not typically produce pellets, and this method is not suitable for those species. Furthermore, this technique does not allow researchers to quantify how much plastic is left in the bird—unlike lavage, emetics, or gastrointestinal analysis of carcasses. As this technique is used with a variety of species, it is important to consider species-specific differences in the amount of plastic ejected in pellets and retained in the bird.
Fig. 3.

Different types of pellets have different production rates (Votier et al. 2001). Single pellets may not represent single meals, but an approximate conversion rate can be calculated based on the species of interest (Ryan and Fraser 1988; Votier et al. 2001; Hammer et al. 2016). In contrast to the accumulated plastics found during stomach dissections, pellets, like regurgitations, represent a snapshot of only a short period of time. Most pellet collections are also limited to the breeding season when marine birds are at terrestrial nesting sites, limiting seasonal comparisons in many species.
When collecting pellets, their condition should be graded. The type of prey that constitute the pellets may determine how long they will last on the ground before they disintegrate, which may bias the amount of plastic found in one type of pellet over another. To overcome this, the integrity of the pellet should be recorded as: (1) fresh/wet (not to be mistaken for wet due to precipitation) and structurally intact, (2) dry and structurally intact, (3) partially broken down, and (4) completely broken down (individual fragments found). The likelihood of finding plastic decreases as the integrity of the pellet decreases, and it is impossible to associate any plastic found with completely broken down pellets. In some species, like the flesh-footed shearwater (Ardenna carneipes (Gould, 1844)), chicks often cast a pellet when they emerge from their burrow to fledge (Lavers et al. 2014). These pellets are not often well defined, but instead appear more like a “spray” of plastic on the ground (Fig. 4). Unlike pellets from gulls and skuas, those from shearwaters are more difficult to identify and delineate, particularly when they contain pieces of pumice, which resembles the colony substrate (Fig. 5). Ideally, all pellets should be individually stored in clean bags or jars. If this is not possible, pellets can be stored based on their decomposition status, with intact pellets bagged together, and all partially or completely degraded pellets bagged individually.
Fig. 4.

Fig. 5.

Processing methods
Necropsies
The purposes of necropsy are to determine cause of death, collect data about the bird (morphometrics, condition, etc.), and collect the gastrointestinal tract (GIT) and any tissues of interest. Regardless of the source, all carcasses destined for plastics examination should be handled carefully to ensure that the GIT is complete upon examination (Fig. 2). If carcasses have been opened by decomposition or scavengers, ingested debris assessments may be either incomplete due to pieces being lost to the environment or inflated because of additional plastic pieces from the environment. The GIT may be preserved (e.g., freezing or submersion in ethanol) so that it can be processed under controlled laboratory conditions at a later date.
When examining GITs, the appropriate location within the GIT must be identified and noted explicitly (Fig. 2). Although “GIT” is often used when referring to ingested debris in seabirds, often only portions of the GIT are examined (Fig. 2). Most seabird studies examine the gizzard and proventriculus (van Franeker et al. 2011), some studies report separate values for these two compartments (Mallory 2008), and others report plastic burdens from the entire length of GIT, including the intestines (Provencher et al. 2010). This often varies due to the original purpose of the study. For plastic accumulation studies, the gizzard and proventriculus are where plastics are retained. Plastic or debris items found in the intestines are likely small enough to be passed through the GIT, and will be excreted by the individual (Provencher et al. 2018).
When possible, debris contents for different sections of the GIT should be recorded separately. In monitoring of northern fulmars, contents of the proventriculus and gizzard (Fig. 2) are combined for cost efficiency of the monitoring program (OSPAR 2017), but in the parallel monitoring of sea turtles in other areas of the European Union, esophagus, stomach, and gut contents are all quantified separately as these different GIT sections all regularly contain plastics in highly variable quantities within and among species, which may be an issue for separate reporting in the future (Matiddi et al. 2017). This is important to consider within the context of birds as well, because of physiological differences among species and detection differences among sampling techniques. Ideally, first descriptions of plastic ingestion in any species will report plastic loads for each GIT compartment separately, but to ensure comparability of results across time and space, plastic ingestion data for monitoring species should conform to established procedures relevant to the monitoring program.
The GIT walls should also be examined during the dissection. All compartments of the GIT should be flushed with water while manipulating the GIT to ensure that all cracks and crevices are thoroughly washed. This also allows for examination of any embedded litter or plastic in the wall or lining of the GIT (e.g., Seif et al. 2018), or the presence of other non-food items such as pumice. Reporting such occurrences may help to document the possible effects of both plastics and other debris items. Often regular tap water is sufficient as long as the purpose of the study is not focused on microplastics that could contaminate the sample (Carrington 2017). When assessing microplastics (size class <5 mm), a clean laboratory setup is necessary during the dissection to avoid contamination of GIT samples by airborne microfiber pollution (Torre et al. 2016). This includes working surfaces that are easily cleaned and maintained, and proper personal protective equipment used at all times in the lab to prevent cross-contamination of plastics from lab personnel.
Regurgitations from birds
Unlike carcasses, regurgitation samples require less processing between collection and examination for plastics. Preservation of the sample is necessary to minimize decomposition of likely associated food items in the samples if diet questions are of interest as well. The entire regurgitation (solids and liquids) should be stored in a clean container and preserved in alcohol or frozen prior to laboratory examination.
Pellet examination
Unlike for carcasses, little processing is needed for pellets between field collection and plastic examination. After pellets have been collected in the field they should be dried as quickly as possible to prevent fungal growth, and stored in a dry and dark condition before dissection. Samples should be dried either at room temperature or in an oven at 55 °C for 48–72 h. Pellets should be weighed prior to drying and then during the drying process; drying is complete when the pellet has a constant mass. The pellets can then be stored in a dry place until examined for plastic.
Cross-cutting ingested plastics assessment methods
Separation of liquids and solids
For all samples that need to separate solids from liquids (e.g., gut contents from necropsies and regurgitations), a series of stainless steel or brass sieves can be used. Stacked sieves (e.g., 5, 1, and 0.3 mm) can be used to separate different fractions. The 5 mm sieve splits meso/macro plastics (>5 mm) from microplastics (<5 mm). Further sieve sizes will depend on the plastic size of interest, but for most birds a 1 mm sieve should be used as pieces smaller than this are likely passed along through the pyloric sphincter and do not accumulate within the bird. A smaller sieve size (e.g., 0.3 mm) can be used when particularly small species are being examined (e.g., storm-petrels). Most microplastic studies based on net sampling at sea use a mesh size of 0.3 mm. Importantly, if microplastics are of interest special laboratory set-ups are needed to assess microplastics in these smaller size classes accurately (see Examining samples for <1 mm plastics, below). If diet composition is also of interest, this method can also be used to separate prey items. In most cases, once the plastics have been rinsed using sieves the liquid can be discarded. All items in the sieve can be examined. Once diet items of interest are removed, the sample can be further processed to remove any biological material from the sample (see the Removal of organic matter, below).
Removal of sediment
For species that consume shelled organisms, large amounts of sand and hard shells may be present in their guts (e.g., bottom feeding seaducks). Once ingested items are ready to be processed, density separation can be used to separate sand and grit from plastic items and prey items. This can include density separation of solids using a simple 5 mol/L NaCl (density = 1.15 g/mL) solution. Without introducing additional chemicals, the saturated NaCl solution will separate the most common plastic polymers found in seabirds (i.e., polyethylene (d = 0.91–0.97 g/mL), polypropylene (d = 0.94 g/mL), and polystyrene (d = 1.05 g/mL)) from sand and grit. To obtain a higher certainty that all polymer particles will float (e.g., including polyvinyl chloride d = 1.4 g/mL), other solutions may be needed. For example, an aqueous 5.4 mol/L lithium metatungstate solution (d = 1.6 g/mL) may be used (Masura et al. 2015), but these have the risk of introducing unwanted chemicals to the plastic material and are not recommended in cases where the chemical analysis of the ingested plastics are of interest without further testing of how these methods may affect the chemical composition of plastics. After density separation, floating items can be collected using sieves as appropriate. If needed, additional techniques to separate organic matter from plastics can be applied (see the next section).
Removal of organic matter
If samples are cluttered with biological material, a digestive treatment may be needed to separate plastics. This may be the case if carcasses are collected when the birds are feeding, or regurgitations are collected when adults are returning to colonies with large quantities of prey to feed to their chicks. Most digestion protocols have been developed to study microplastic ingestion by fish, invertebrates, and marine birds (Besseling et al. 2015; Courtene-Jones et al. 2017; Kühn et al. 2017).
Digestion protocols consist of four main groups: acidic, alkaline, oxidizing, and enzymatic dissolution (Dehaut et al. 2016). Acidic solutions tend to dissolve some polymer types (Dehaut et al. 2016; Lusher et al. 2017), and may degrade plastics, which can reduce their mass or lead to yellowing (which is important to note in relation to reporting the colour of plastics; Dehaut et al. 2016). Potassium hydroxide (KOH) in 1 mol/L concentrations with heat (60 °C) is generally preferred as the least destructive treatment to the plastics (except cellulose acetate), but experimentation is still needed under a range of conditions and tissue types (Kühn et al. 2017). Enzymatic digestion can also be used for digestion of organic material in samples. Enzymes such as trypsin, papain, and collagenase (Courtene-Jones et al. 2017), and off-the-shelf mixtures such as enzymatic washing detergent (Bravo Rebolledo et al. 2013; Besseling et al. 2015) have been tested on mussels, seal scat, and cetacean intestines. Proteinase-K has also been effectively used to isolate microplastics from biotic samples (Cole et al. 2015; Löder et al. 2017). Cole et al. (2015) found that >97% of the biological material from a sample could be reduced using this technique. Even though the effects of enzymes on plastics have been observed, enzymatic approaches required less time and effort (Lusher et al. 2017). Importantly, however, the concentrations required and the saturation point of enzymatic activity have not been determined for many tissues and sample conditions.
Digestion protocols should only be used when necessary, and the solution and potential impact on the integrity of the sample (i.e., impacts on colour, mass, or degrading of certain polymer types) should be reported. Typically, plastics >1 mm can be identified without digestion protocols by an experienced observer. However, a growing interest in quantifying plastics <1 mm might lead to future research questions regarding smaller plastic particles in seabirds.
Alternative to separation techniques
Where samples are likely to contain microplastic pieces and have high amounts of sediment or biological material that cannot be removed easily, there are approaches such as staining with Nile Red that may be useful. This approach uses the selective staining properties of Nile Red onto plastics surfaces, which then makes plastic pieces fluoresce under blue light (Maes et al. 2017). This technique has been employed in environmental samples, but not in seabird studies to date. This may be a particularly useful method where large sample sizes prohibit detailed examination of the samples as it can be done quickly. The disadvantage of this technique is that it only works to detect plastics, not other debris types, which could be of interest in some studies (Seif et al. 2018). It may also stain the plastics, which may be a disadvantage if colour is a metric of interest.
Drying plastics
Once plastics and other debris items are removed from the sample, all plastics and other items should be thoroughly cleaned and dried before weighing. It is important to note that aggressive rinsing or scrubbing could remove adhered pollutants that may also be of interest in the study; therefore, the goals of the study will determine how well debris items should be rinsed or scrubbed. All debris should be air dried for at least 24 h at room temperature or for a minimum of 12 h in a drying oven at 40 °C on a covered stainless steel or glass plate to prevent any cross-contamination. If porous or foam-based material is involved, drying may require more time. Once plastics are dry, pieces should be weighed individually using an analytical balance, ideally to 0.001 g precision.
Categorization of ingested plastics and other debris
Suggested best practices for categorizing debris and reporting results have been detailed by Provencher et al. (2017).
Other aspects to consider in plastics studies of seabirds
Polymer identification
In addition to visual examination, Raman spectrometry or Fourier-transform infrared spectroscopy (FTIR) can be used to identify the polymer types of plastic (Avery-Gomm et al. 2016). Such polymer identification may also be important when assessing which plastic-associated contaminants may be absorbed by wildlife as a result of accumulated plastics in the gut (Rochman et al. 2014; Koelmans et al. 2016).
Some of the digestion procedures outlined above may alter the chemical composition of some polymers, leading to challenges in accurate polymer identification. For example, the alkaline solution NaOH degraded cellulose carbonate, polycarbonate, and polyethylene terephthalate (Karami et al. 2017), whereas the oxidizing peroxide sulfate potassium did not harm polymer structure but was less efficient in dissolving organic material and had some handling issues that were of concern (Karami et al. 2017).
Contaminant analysis of plastics
For some research questions it may be useful to analyze the ingested plastics themselves for contaminants (Rochman et al. 2013). If ingested plastics are to be analyzed for intrinsic and adsorbed contaminants, separation of the plastics from organic matter may need to be done manually to ensure that the contaminant concentrations are those as they were found in the stomachs and not altered due to a chemical reaction in the sorting process. Plastics should then also be kept in chemically clean containers (rinsed thoroughly with a solvent) between dissection and chemical analysis (Rochman et al. 2013).
Examining samples for <1 mm plastics
Regurgitated pellets are not appropriate to quantify ingested microfibers and other small plastics because of the high levels of environmental contamination. Plastic pieces that are <1 mm do not likely accumulate in most seabirds, and therefore seabirds are not likely to be useful indicators of the distribution of this small microplastic pollution in the environment. However, there may be research questions related to the plastic excretion by seabirds, or evaluating seabirds as vectors for plastics pollution through their guano (van Franeker 2011; Provencher et al. 2018).
If samples are being used to determine the ingestion of plastics <1 mm, there are cross-contamination procedures that must be put into practice to ensure that rigorous and robust findings are reported (Torre et al. 2016; Hermsen et al. 2017; Wesch et al. 2017). Samples are at risk from contamination by particles present in re-used sample containers, on the clothes of workers, airborne (clothing) fibres, and (or) by improperly sealed samples (Torre et al. 2016; Wesch et al. 2017). The use of filtered water may be necessary given that tap water may introduce microplastics to a sample (Vermaire et al. 2017). Personal protection equipment such as laboratory coats, gloves, and even hairnets or hats may be necessary to reduce fibres from lab workers from entering the sample during the entire dissection process (Torre et al. 2016). Protective clothing that is a bright or uncommon colour (i.e., pink or orange) can be used, as microfibers of these colours can then be discounted from the final tally of debris in a sample.
Isolated spaces where plastics <1 mm will be examined can be effective at reducing cross-contamination. Creating an isolated stereomicroscope area for visual identification can reduce airborne contamination (Torre et al. 2016), and is commonly used in microplastic studies in other biota (Davidson and Dudas 2016). Sample blanks must be used to detect and correct for any cross-contamination that does occur (Vermaire et al. 2017). This can be done through opening a set of empty jars for the same amount of time for which a standard seabird sample is examined, and then using the same quantification techniques. If digestion of the samples is done, similarly clean jars should be subjected to the same processes and then examined. The maximum number of microfibers in the blanks can then be subtracted from each sample as a conservative approach to accounting for such cross-contamination (Vermaire et al. 2017; Provencher et al. 2018). On a smaller more portable scale, the use of glove boxes that create a confined air space around samples can be used to reduce environmental microfibers from being introduced into the samples (Torre et al. 2016). These methods may not be applicable to most seabird studies, which focus on plastics >1 mm, but are useful when examining microplastics.
Data processing and presentation
Variables related to debris ingestion to be reported
Many factors influence metrics of ingestion of marine debris in seabirds, including species, sex, age, and breeding stage. If these metrics are not of direct interest to the published manuscript containing the plastic ingestion data, they should still be recorded for each sample (e.g., each carcass) and reported in supplementary online material or in open data archives as they may be critical to future studies and comparisons (e.g., O’Hanlon et al. 2017).
Reporting of ingested plastic metrics in seabirds
Standardized reporting recommendations have been discussed extensively by Provencher et al. (2017). Briefly, as a minimum, the location of sampling, the type of sampling, sample size, and the frequency of occurrence of ingested plastics should be reported. If specific types of plastic items can be identified, they should be reported. To allow for a full understanding of the data, the mean (with standard deviation and error), median, and range of mass of ingested plastics per individual, including all individuals sampled, and the mean (with standard deviation and error), median, and range of all plastics reported by debris category (see above) should be reported. In the context of reporting microplastics <1 mm, cross-contamination cannot be excluded in most cases (Wesch et al. 2017); therefore, fibres should always be reported separately and all measures taken to prevent contamination should be mentioned.
Although documenting and reporting ingested plastics is critical to understanding global patterns, it is also critical to use these standardized approaches when reporting “zeroes”; this is analogous to issues arising from meta-analyses and the importance of publishing non-significant results (Koricheva 2003; Dwan et al. 2013). When standardized protocols are applied and no plastics are found, these data should be published. Establishing the absence of plastics in species or individuals will enhance our understanding of the pervasiveness of plastic pollution, the potential impacts of plastic ingestion, and allow for the development of more robust predictive models for data-limited species.
Discussion
We detailed standardized approaches for the three main techniques for evaluating plastic ingestion by seabirds (necropsies, regurgitations, and pellets), including field collection, laboratory processing and data presentation. Creating a single protocol that will be widely applicable to hundreds of species from the emperor penguin (Aptenodytes forsteri Gray, 1844; 23 kg) to the least storm-petrel (Halocyptena microsoma (Coues, 1864); 25 g). Therefore, we have purposely presented an overview of harmonized best practices for assessing ingested plastics in marine birds and not a detailed protocol. There is room for the development of specific, step-by-step protocols for groups of species, or approaches that are used across a suite of sampling techniques, and we hope that this discussion will inform future efforts.
Given the breadth of plastic pollution information that can and has been gleaned from seabird studies, there is an immediate need to move beyond simply reporting plastic and litter ingestion by birds to allow the field to address larger, ecologically relevant questions. As any field of research matures, adoption of standardized methods becomes important. Moving forward, plastic ingestion studies employing standardized methods for sample collection, processing, plastic quantification, and reporting will provide valuable data points that will make statistically and scientifically credible large-scale comparisons possible (Stewart 2010). This will enhance our understanding of the distribution of plastics in the environment, which species are most vulnerable to plastic ingestion, and the potential population-level impacts of plastic ingestion.
Acknowledgements
We thank all those researchers who continue to work on plastic ingestion by seabirds and report those findings in the peer-reviewed literature. This paper was produced as a contribution of the Plastics Working Group of the World Seabird Union. JFP was supported by a G. Weston Foundation Post-doctoral in Northern Research Fellowship. SBB is supported by the David H. Smith Fellowship. ALB and JLL are supported by BirdLife Tasmania, Detached Foundation (P. Clive and B. Neill), Save our Shearwaters, Sea World Research and Rescue Foundation (SWR/4/2015), Sydney Sea Life Foundation, Trading Consultants Ltd. (V. Wellington), L. Bryce, the W.V. Scott Charitable Trust, and numerous private donors. SK conducts her Ph.D. plastics research at Wageningen Marine Research under the PLASTOX project, funded by the Earth and Life Sciences section of the Netherlands Organisation for Scientific Research (ALW-NWO project 856.15.001) under the umbrella of the EU JPI-Oceans program. SH is funded by the Faroese Research Council and Statoil Faroes. SAG is supported by funding from the University of Queensland and the National Scientific and Engineering Research Council of Canada. MLM was supported by a Natural Sciences and Engineering Research Council grant, as well as a Chair position from Fulbright Canada. Lastly, we thank the three anonymous reviewers who contributed constructive comments on this manuscript that helped to improve the work.
References
Acampora H, Schuyler QA, Townsend KA, and Hardesty BD. 2014. Comparing plastic ingestion in juvenile and adult stranded short-tailed shearwaters (Puffinus tenuirostris) in eastern Australia. Marine Pollution Bulletin, 78(1–2): 63–68.
Álvarez G, Barros Á, and Velando A. 2018. The use of European shag pellets as indicators of microplastic fibers in the marine environment. Marine Pollution Bulletin, 137: 444–448.
Auman HJ, Ludwig JP, Giesy JP, and Colborn T. 1998. Plastic ingestion by Laysan albatross chicks on Sand Island, Midway Atoll, in 1994 and 1995. In Albatross: biology and conservation. Edited by G Robertson and R Gales. Surrey Beatty & Sons Pty Limited, Chipping Norton, New South Wales, Australia. pp. 239–244.
Avery-Gomm S, O’Hara PD, Kleine L, Bowes V, Wilson LK, and Barry KL. 2012. Northern fulmars as biological monitors of trends of plastic pollution in the eastern North Pacific. Marine Pollution Bulletin, 64(9): 1776–1781.
Avery-Gomm S, Provencher JF, Morgan KH, and Bertram DF. 2013. Plastic ingestion in marine-associated bird species from the eastern North Pacific. Marine Pollution Bulletin, 72(1): 257–259.
Avery-Gomm S, Valliant M, Schacter CR, Robbins KR, Liboiron M, Daoust PY, et al. 2016. A study of wrecked Dovekies (Alle alle) in the western North Atlantic highlights the importance of using standardized methods to quantify plastic ingestion. Marine Pollution Bulletin, 113(1–2): 75–80.
Avery-Gomm S, Provencher JF, Liboiron M, Poon FE, and Smith PA. 2018. Plastic pollution in the Labrador Sea: an assessment using the seabird northern fulmar Fulmarus glacialis as a biological monitoring species. Marine Pollution Bulletin, 127: 817–822.
Barrett RT, Camphuysen K, Anker-Nilssen T, Chardine JW, Furness RW, Garthe S, et al. 2007. Diet studies of seabirds: a review and recommendations. ICES Journal of Marine Science, 64(9): 1675–1691.
Besseling E, Foekema EM, van Franeker JA, Leopold MF, Kühn S, Bravo Rebolledo EL, et al. 2015. Microplastic in a macro filter feeder: humpback whale Megaptera novaeangliae. Marine Pollution Bulletin, 95(1): 248–252.
Boesman P, Bonan A, Chernasky A, Christie D, Collar N, Copete JL, et al. 2017. Handbook of the birds of the world. HBW Alive [online]: Available from hbw.com/.
Bond AL, and Lavers JL. 2013. Effectiveness of emetics to study plastic ingestion by Leach’s Storm-petrels (Oceanodroma leucorhoa). Marine Pollution Bulletin, 70: 171–175.
Bond AL, Jones IL, Williams JC, and Byrd GV. 2010. Auklet (Charadriiformes: Alcidae, Aethia spp.) chick meals from the Aleutian Islands, Alaska, have a very low incidence of plastic marine debris. Marine Pollution Bulletin, 60(8): 1346–1349.
Borrelle SB, Rochman CM, Liboiron M, Bond AL, Lusher A, Bradshaw H, et al. 2017. Opinion: why we need an international agreement on marine plastic pollution. Proceedings of the National Academy of Sciences of the USA, 114(38): 9994–9997.
Bravo Rebolledo EL, Van Franeker JA, Jansen OE, and Brasseur SMJM. 2013. Plastic ingestion by harbour seals (Phoca vitulina) in The Netherlands. Marine Pollution Bulletin, 67(1–2): 200–202.
Cadée GC. 2002. Seabirds and floating plastic debris. Marine Pollution Bulletin, 44(11): 1294–1295.
Carrington D. 2017. Plastic fibres found in tap water around the world, study reveals [online]: Available from theguardian.com/environment/2017/sep/06/plastic-fibres-found-tap-water-around-world-study-reveals.
Cole M, Webb H, Lindeque PK, Fileman ES, Halsband C, and Galloway TS. 2015. Isolation of microplastics in biota-rich seawater samples and marine organisms. Scientific Reports, 4(1): 4528.
Courtene-Jones W, Quinn B, Murphy F, Gary SF, and Narayanaswamy BE. 2017. Optimisation of enzymatic digestion and validation of specimen preservation methods for the analysis of ingested microplastics. Analytical Methods, 9(9): 1437–1445.
Davidson K, and Dudas SEE. 2016. Microplastic ingestion by wild and cultured Manila clams (Venerupis philippinarum) from Baynes Sound, British Columbia. Archives of Environmental Contamination and Toxicology, 71(2): 147–156.
Dehaut A, Cassone A-L, Frère L, Hermabessiere L, Himber C, Rinnert E, et al. 2016. Microplastics in seafood: benchmark protocol for their extraction and characterization. Environmental Pollution, 215: 223–233.
Donnelly-Greenan EL, Harvey JT, Nevins HM, Hester MM, and Walker WA. 2014. Prey and plastic ingestion of Pacific Northern Fulmars (Fulmarus glacialis rogersii) from Monterey Bay, California. Marine Pollution Bulletin, 85(1): 214–224.
Dwan K, Gamble C, Williamson PR, and Kirkham JJ. 2013. Systematic review of the empirical evidence of study publication bias and outcome reporting bias—an updated review. PLoS ONE, 8(7): e66844.
English MD, Robertson GJ, Avery-Gomm S, Pirie-Hay D, Roul S, Ryan PC, et al. 2015. Plastic and metal ingestion in three species of coastal waterfowl wintering in Atlantic Canada. Marine Pollution Bulletin, 98(1–2): 349–353.
Fife DT, Robertson GJ, Shutler D, Braune BM, and Mallory ML. 2015. Trace elements and ingested plastic debris in wintering dovekies (Alle alle). Marine Pollution Bulletin, 91(1): 368–371.
Fijn RC, van Franeker JA, and Trathan PN. 2012. Dietary variation in chick-feeding and self-provisioning Cape Petrel Daption capense and Snow Petrel Pagodroma nivea at Signy Island, South Orkney Islands, Antarctica. Marine Ornithology, 40(2): 81–87.
Fry DM, Fefer SI, and Sileo L. 1987. Ingestion of plastic debris by Laysan Albatrosses and Wedge-tailed Shearwaters in the Hawaiian Islands. Marine Pollution Bulletin, 18(6 Suppl. B): 339–343.
Furness RW. 1985. Plastic particle pollution: accumulation by procellariiform seabirds at Scottish Colonies. Marine Pollution Bulletin, 16(3): 103–106.
Gaston AJ. 2004. Seabirds: a natural history. Yale University Press, New Haven, Connecticut.
Gaunt AS, and Oring LW. 1997. Guidelines to the use of wild birds in research. Washington, D.C.
Gil-Delgado JA, Guijarro D, Gosálvez RU, López-Iborra GM, Ponz A, and Velasco A. 2017. Presence of plastic particles in waterbirds faeces collected in Spanish lakes. Environmental Pollution, 220: 732–736.
Goldsworthy B, Young MJ, Seddon PJ, and van Heezik Y. 2016. Stomach flushing does not affect apparent adult survival, chick hatching, or fledging success in yellow-eyed penguins (Megadyptes antipodes). Biological Conservation, 196: 115–123.
Hammer S, Nager RG, Johnson PCD, Furness RW, and Provencher JF. 2016. Plastic debris in great skua (Stercorarius skua) pellets corresponds to seabird prey species. Marine Pollution Bulletin, 103(1–2): 206–210.
Harper PC, and Fowler JA. 1987. Plastic pellets in New Zealand storm-killed prions (Pachyptila spp.) 1958–1977. Notornis, 34(1): 65–70.
Hermsen E, Pompe R, Besseling E, and Koelmans AA. 2017. Detection of low numbers of microplastics in North Sea fish using strict quality assurance criteria. Marine Pollution Bulletin, 122(1–2): 253–258.
Herzke D, Anker-Nilssen T, Nøst TH, Götsch A, Christensen-Dalsgaard S, Langset M, et al. 2016. Negligible impact of ingested microplastics on tissue concentrations of persistent organic pollutants in northern fulmars off coastal Norway. Environmental Science & Technology, 50(4): 1924–1933.
Holland ER, Mallory ML, and Shutler D. 2016. Plastics and other anthropogenic debris in freshwater birds from Canada. Science of the Total Environment, 571: 251–258.
Hunt GL Jr. 1972. Influence of food distribution and human disturbance on the reproductive success of herring gulls. Ecology, 53: 1051–1061.
Hutton I, Carlile N, and Priddel D. 2008. Plastic ingestion by Flesh-footed Shearwaters, Puffinus carneipes, and Wedge-tailed Shearwaters, Puffinus pacificus. Papers and Proceedings of the Royal Society of Tasmania, 142(1): 67–72.
Jahncke J, Garcia-Godos A, and Goya E. 1999. The diet of the Peruvian Diving-Petrel at La Vieja and San Gallan, Peru. Journal of Field Ornithology, 70(1): 71–79.
Janinhoff N, Verdaat H, and van Franeker JA. 2010. Rosse Franjepoot Phalaropus fulicaria fourageert in plastic soep [Grey phalarope foraging in plastic soup]. Sula, 23(1): 41–45.
Jiménez S, Domingo A, Brazeiro A, Defeo O, and Phillips RA. 2015. Marine debris ingestion by albatrosses in the southwest Atlantic Ocean. Marine Pollution Bulletin, 96(1–2): 149–154.
Karami A, Golieskardi A, Choo CK, Romano N, Ho YB, and Salamatinia B. 2017. A high-performance protocol for extraction of microplastics in fish. Science of the Total Environment, 578: 485–494.
Kenyon KW, and Kridler E. 1969. Laysan Albatrosses swallow indigestible matter. Auk, 86(2): 339–343.
Koelmans AA, Bakir A, Burton GA, and Janssen CR. 2016. Microplastic as a vector for chemicals in the aquatic environment: critical review and model-supported reinterpretation of empirical studies. Environmental Science & Technology, 50(7): 3315–3326.
Koricheva J. 2003. Non-significant results in ecology: a burden or a blessing in disguise? Oikos, 102(2): 397–401.
Kühn S, Bravo Rebolledo EL, and van Franeker JA. 2015. Deleterious effects of litter on marine life. In Marine anthropogenic litter. Edited by M Bergmann, L Gutow, and M Klages. Springer Open, Bremerhaven, Germany. pp. 75–116.
Kühn S, van Werven B, van Oyen A, Meijboom A, Bravo Rebolledo EL, and van Franeker JA. 2017. The use of potassium hydroxide (KOH) solution as a suitable approach to isolate plastics ingested by marine organisms. Marine Pollution Bulletin, 115(1–2): 86–90.
Lavers JL, Bond AL, and Hutton I. 2014. Plastic ingestion by Flesh-footed Shearwaters (Puffinus carneipes): implications for fledgling body condition and the accumulation of plastic-derived chemicals. Environmental Pollution, 187: 124–129.
Law KL, Morét-Ferguson S, Maximenko NA, Proskurowski G, Peacock EE, Hafner J, et al. 2010. Plastic accumulation in the North Atlantic subtropical gyre. Science, 329(5996): 1185–1188.
Lindborg VA, Ledbetter JF, Walat JM, and Moffett C. 2012. Plastic consumption and diet of Glaucous-winged Gulls (Larus glaucescens). Marine Pollution Bulletin, 64(11): 2351–2356.
Löder MGJ, Imhof HK, Ladehoff M, Löschel LA, Lorenz C, Mintenig S, et al. 2017. Enzymatic purification of microplastics in environmental samples. Environmental Science & Technology, 51(24): 14283–14292.
Lusher ALL, Welden NAA, Sobral P, and Cole M. 2017. Sampling, isolating and identifying microplastics ingested by fish and invertebrates. Analytical Methods, 9(9): 1346–1360.
Maes T, Jessop R, Wellner N, Haupt K, and Mayes AG. 2017. A rapid-screening approach to detect and quantify microplastics based on fluorescent tagging with Nile Red. Scientific Reports, 7: 44501.
Mallory ML. 2008. Marine plastic debris in northern fulmars from the Canadian high Arctic. Marine Pollution Bulletin, 56(8): 1501–1504.
Masura J, Baker J, Foster G, and Arthur C. 2015. Laboratory methods for the analysis of microplastics in the marine environment: recommendations for quantifying synthetic particles in waters and sediments. NOAA, Seattle, Washington.
Matiddi M, Hochsheid S, Camedda A, Baini M, Cocumelli C, Serena F, et al. 2017. Loggerhead sea turtles (Caretta caretta): a target species for monitoring litter ingested by marine organisms in the Mediterranean Sea. Environmental Pollution, 230: 199–209.
Matthews LH. 1949. The origin of stomach oil in the petrels, with comparative observations on the avian proventriculus. Ibis, 91(3): 373–392.
O’Hanlon NJ, James NA, Masden EA, and Bond AL. 2017. Seabirds and marine plastic debris in the northeastern Atlantic: a synthesis and recommendations for monitoring and research. Environmental Pollution, 231: 1291–1301.
OSPAR. 2015. Guidelines for monitoring and assessment of plastic particles in stomachs of fulmars in the North Sea area. In OSPAR Commission Agreement 2015-03 (Source: EIHA 15/5/12 Add.1). OSPAR, Texel, the Netherlands [online]: Available from ospar.org/convention/agreements?q=fulmar&t=32281&a=&s=.
OSPAR. 2017. OSPAR Intermediate Assessment 2017—plastic particles in fulmar stomachs in the North Sea. OSPAR Assessment Portal OAP [online]: Available from oap-cloudfront.ospar.org/media/filer_public/f4/34/f434a292-3fad-466c-9e89-c2ff09dc9e9d/fulmar.pdf.
Provencher JF, Gaston AJ, Mallory ML, O’Hara PD, and Gilchrist HG. 2010. Ingested plastic in a diving seabird, the thick-billed murre (Uria lomvia), in the eastern Canadian Arctic. Marine Pollution Bulletin, 60(9): 1406–1411.
Provencher JF, Bond AL, Hedd A, Montevecchi WA, Muzaffar SB, Courchesne SJ, et al. 2014. Prevalence of marine debris in marine birds from the North Atlantic. Marine Pollution Bulletin, 84(1–2): 411–417.
Provencher JF, Bond AL, Avery-Gomm S, Borrelle SB, Bravo Rebolledo EL, Hammer S, et al. 2017. Quantifying ingested debris in marine megafauna: a review and recommendations for standardization. Analytical Methods, 9(9): 1454–1469.
Provencher JF, Vermaire JC, Avery-Gomm S, Braune BM, and Mallory ML. 2018. Garbage in guano? Microplastic debris found in faecal precursors of seabirds known to ingest plastics. Science of the Total Environment, 644: 1477–1484.
Reisser J, Slat B, Noble K, du Plessis K, Epp M, Proietti M, et al. 2015. The vertical distribution of buoyant plastics at sea: an observational study in the North Atlantic Gyre. Biogeosciences, 12(4): 1249–1256.
Robertson G, Kent S, and Seddon J. 1980. Effects of the water-offloading technique on Adélie Penguins. Journal of Field Ornithology, 65(3): 376–380.
Rochman CM, Hoh E, Hentschel BT, and Kaye S. 2013. Long-term field measurement of sorption of organic contaminants to five types of plastic pellets: implications for plastic marine debris. Environmental Science & Technology, 47: 1646–1654.
Rochman CM, Hentschel BT, and Teh SJ. 2014. Long-term sorption of metals is similar among plastic types: implications for plastic debris in aquatic environments. PLoS ONE, 9(1): e85433.
Rodríguez A, Ramírez F, Carrasco MN, and Chiaradia A. 2018. Seabird plastic ingestion differs among collection methods: examples from the short-tailed shearwater. Environmental Pollution, 243: 1750–1757.
Roman L, Schuyler QA, Hardesty BD, and Townsend KA. 2016. Anthropogenic debris ingestion by avifauna in eastern Australia. PLoS ONE, 11(8): e0158343.
Ryan PG, and Fraser MW. 1988. The use of great skua pellets as indicators of plastic pollution in seabirds. Emu—Austral Ornithology, 88: 16–19.
Ryan PG, and Jackson S. 1986. Stomach pumping—is killing seabirds necessary? Auk, 103(2): 427–428.
Seif S, Provencher JF, Avery-Gomm S, Daoust P-Y, Mallory ML, and Smith PA. 2018. Plastic and non-plastic debris ingestion in three gull species feeding in an urban landfill environment. Archives of Environmental Contamination and Toxicology, 74(3): 349–360.
Song YK, Hong SH, Jang M, Kang J-H, Kwon OY, Han GM, et al. 2014. Large accumulation of micro-sized synthetic polymer particles in the sea surface microlayer. Environmental Science & Technology, 48(16): 9014–9021.
Stewart G. 2010. Meta-analysis in applied ecology. Biology Letters, 6(1): 78–81.
Threlfall W. 1968. The food of three species of gulls in Newfoundland. Canadian Field-Naturalist, 82: 176–180.
Torre M, Digka N, Anastasopoulou A, Tsangaris C, and Mytilineou C. 2016. Anthropogenic microfibres pollution in marine biota. A new and simple methodology to minimize airborne contamination. Marine Pollution Bulletin, 113(1–2): 55–61.
van Franeker JA. 2011. Reshape and relocate: seabirds as transformers and transporters of microplastics. In Fifth International Marine Debris Conference, Honolulu, Hawaii, 20–25 March 2011.
van Franeker JA, and Law KL. 2015. Seabirds, gyres and global trends in plastic pollution. Environmental Pollution, 203: 89–96.
van Franeker JA, and Meijboom A. 2002. Litter NSV—marine litter monitoring by northern fulmars: a pilot study. Alterra, Wageningen, the Netherlands.
van Franeker JA, Williams R, Imber MJ, and Wolff WJ. 2001. Diet and foraging ecology of Southern Fulmar Fulmarus glacialoides, Antarctic Petrel Thalassoica antarctica, Cape Petrel Daption capense and Snow Petrels Pagodroma nivea ssp. on Ardery Island, Wilkes Land, Antarctica. Ph.D. dissertation, University of Groningen, Texel, the Netherlands.
van Franeker JA, Blaize C, Danielsen J, Fairclough K, Gollan J, Guse N, et al. 2011. Monitoring plastic ingestion by the northern fulmar Fulmarus glacialis in the North Sea. Environmental Pollution, 159(10): 2609–2615.
Vermaire JC, Pomeroy C, Herczegh SM, Haggart O, and Murphy M. 2017. Microplastic abundance and distribution in the open water and sediment of the Ottawa River, Canada, and its tributaries. FACETS, 2: 301–314.
Votier SC, Bearhop S, Ratcliffe N, and Furness RW. 2001. Pellets as indicators of diet in great skuas Catharacta skua. Bird Study, 48: 373–376.
Wesch C, Elert AM, Wörner M, Braun U, Klein R, and Paulus M. 2017. Assuring quality in microplastic monitoring: about the value of clean-air devices as essentials for verified data. Scientific Reports, 7(1): 5424.
Wilson LJ, Daunt F, and Wanless S. 2004. Self-feeding and chick provisioning diet differ in the common guillemot Uria aalge. Ardea, 92(2): 197–207.
Information & Authors
Information
Published In

FACETS
Volume 4 • Number 1 • June 2019
Pages: 111 - 130
Editor: Brett Favaro
History
Received: 29 October 2018
Accepted: 7 January 2019
Version of record online: 9 May 2019
Copyright
© 2019 Provencher et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper.
Key Words
Sections
Subjects
Authors
Author Contributions
All conceived and designed the study.
All performed the experiments/collected the data.
All analyzed and interpreted the data.
All contributed resources.
All drafted or revised the manuscript.
Competing Interests
MLM is currently serving as a Subject Editor for FACETS, but was not involved in review or editorial decisions regarding this manuscript. At the time when he was offered the opportunity to handle the paper the handling editor (Dr. B. Favaro) acknowledged that he and Dr. Provencher are both Liber Ero Fellows and that they know each other, have worked with each other, and are preparing to co-supervise students. Dr. Favaro declared that he was nonetheless able to deliver an unbiased handling of the peer review of the paper to inform the eventual editorial decision.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Jennifer F. Provencher, Stephanie B. Borrelle, Alexander L. Bond, Jennifer L. Lavers, Jan A. van Franeker, Susanne Kühn, Sjúrður Hammer, Stephanie Avery-Gomm, and Mark L. Mallory. 2019. Recommended best practices for plastic and litter ingestion studies in marine birds: Collection, processing, and reporting. FACETS.
4(1): 111-130. https://doi.org/10.1139/facets-2018-0043
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Plastics in biota: technological readiness level of current methodologies
2. Microplastics footprint in nature reserves-a case study on the microplastics in the guano from Yancheng Wetland Rare Birds National Nature Reserve, China
3. Plastic ingestion, accumulated heavy metals, and health metrics of four Larus gull species feeding at a coastal landfill in eastern Canada
4. Among-colony variation in plastic ingestion by Yellow-legged gulls (Larus michahellis) across the western Mediterranean basin
5. Plastic ingestion and trace element contamination of Manx shearwaters Puffinus puffinus on the Faroe Islands
6. An assessment of microplastics in fecal samples from polar bears (Ursus maritimus) in Canada's North
7. Indigo-dyed cellulose fibers and synthetic polymers in surface-feeding seabird chick regurgitates from the Gulf of Alaska
8. The relationship between plastic ingestion and trace element concentrations in Arctic seabirds
9. Microplastic in an apex predator: evidence from Barn owl (Tyto alba) pellets in two sites with different levels of anthropization
10. Birds as bioindicators of plastic pollution in terrestrial and freshwater environments: A 30-year review
11. Geographical and ecological factors affect microplastic body burden in marine fish at global scale
12. Leakage of plastics and other debris from landfills to a highly protected lake by wintering gulls
13. How do life history and behaviour influence plastic ingestion risk in Canadian freshwater and terrestrial birds?
14. Cory's shearwater as a key bioindicator for monitoring floating plastics
15. Identifying laboratory sources of microplastic and nanoplastic contamination from the air, water, and consumables
16. Exploring the pathology of liver, kidney, muscle, and stomach of fledgling seabirds associated with plastic ingestion
17. An assessment of microplastics in fecal samples from polar bears (<i>Ursus maritimus</i>) in Canada’s North
18. First report on occurrence and characterization of microplastics in feces of Corvus splendens (Vieillot, 1817)
19. The challenges of opportunistic sampling when comparing prevalence of plastics in diving seabirds: A multi-species example from Norway
20. Beached seabirds as plastic biomonitors in Brazil from the Beach Monitoring Project of the Santos Basin (PMP-BS)
21. The threat of microplastics: Exploring pollution in coastal ecosystems and migratory shorebirds along the west coast of India
22. Global assessment of marine plastic exposure risk for oceanic birds
23. Microplastics in gastrointestinal tracts of gentoo penguin (Pygoscelis papua) chicks on King George Island, Antarctica
24. No microplastics detected in the first assessment of Atlantic walrus stomachs from Nunavut, Canada
25. Biovectoring of plastic by white storks from a landfill to a complex of salt ponds and marshes
26. Evaluation of meso- and microplastic ingestion by the northern fulmar through a non-lethal sampling method
27. Dietary evidence of trophic segregation between Campbell albatross Thalassarche impavida and grey-headed albatross T. chrysostoma at subantarctic Campbell Island
28. Microplastics in feces and pellets from yellow-legged gull (Larus michahellis) in the Atlantic Islands National Park of Galicia (NW Spain)
29. Marine debris ingestion by adults and fledglings of Swinhoe's storm petrels in the Republic of Korea
30. Assessing plastic ingestion in birds of prey from British Columbia, Canada
31. Guanay cormorant (Leucocarbo bougainvilliorum) pellets as an indicator of marine plastic pollution along the Peruvian coast
32. First Report on Microplastics Quantification in Poultry Chicken and Potential Human Health Risks in Pakistan
33. Current levels of microplastic pollution impact wild seabird gut microbiomes
34. Finding the Balance between Research and Monitoring: When Are Methods Good Enough to Understand Plastic Pollution?
35. Implications of Regurgitative Feeding on Plastic Loads in Northern Fulmars (
Fulmarus glacialis
): A Study from Svalbard
36. Future monitoring of litter and microplastics in the Arctic—challenges, opportunities, and strategies
37. Can the mass of plastic ingested by seabirds be predicted by the number of ingested items?
38. Changes in plastic ingestion by yellow-legged gulls (Larus michahellis) over the breeding season
39. New Methods for the Quantification of Ingested Nano- and Ultrafine Plastics in Seabirds
40. Microplastic in northern anchovies (Engraulis mordax) and common murres (Uria aalge) from the Monterey Bay, California USA - Insights into prevalence, composition, and estrogenic activity
41. Pollution—Lights, plastics, oil, and contaminants
42. High incidence of plastic debris in Andean condors from remote areas: Evidence for marine-terrestrial trophic transfer
43. The one-two punch of plastic exposure: Macro- and micro-plastics induce multi-organ damage in seabirds
44. Shorebirds ingest plastics too: what we know, what we do not know, and what we should do next
45. No evidence of microplastic ingestion in emperor penguin chicks (Aptenodytes forsteri) from the Atka Bay colony (Dronning Maud Land, Antarctica)
46. Microplastic Ingestion Induces Size-Specific Effects in Japanese Quail
47. Ingested plastics in beach-washed Fairy Prions Pachyptila turtur from Tasmania
48. Towards a North Pacific Ocean long-term monitoring program for plastic pollution: A review and recommendations for plastic ingestion bioindicators
49. Macrophage–NLRP3 Activation Promotes Right Ventricle Failure in Pulmonary Arterial Hypertension
50. Far from a distraction: Plastic pollution and the planetary emergency
51. First documentation of plastic ingestion in the arctic glaucous gull (Larus hyperboreus)
52. Future monitoring of litter and microplastics in the Arctic – challenges, opportunities and strategies
53. Microplastics in the Gulf of Mexico: A Bird’s Eye View
54. Comparing methods for monitoring nest debris using silver gulls as a case study
55. The potential of aerial insectivores for monitoring microplastics in terrestrial environments
56. Fatty acids composition in yellow-legged (Larus michahellis) and lesser black-backed (Larus fuscus) gulls from natural and urban habitats in relation to the ingestion of anthropogenic materials
57. Birds of a Feather Eat Plastic Together: High Levels of Plastic Ingestion in Great Shearwater Adults and Juveniles Across Their Annual Migratory Cycle
58. Consequences of Plastic Trash on Behavior and Ecology of Birds
59. Degradation of Plastic Waste in the Marine Environment
60. Current methods and future directions in avian diet analysis
61. Community-scientist collaboration in the creation, management and research for two National Wildlife Areas in Arctic Canada
62. A 6-year survey of plastic ingestion by aquatic birds in southern Portugal
63. Plastic ingestion in adult and fledgling Manx Shearwaters Puffinus puffinus on Skomer Island, Wales
64. Microplastic in fish – A global synthesis
65. Annual plastic ingestion and isotopic niche patterns of two sympatric gull species at Newfoundland, Canada
66. The broader isotopic niche of Long-tailed Duck Clangula hyemalis implies a higher risk of ingesting plastic and non-plastic debris than for other diving seabirds
67. Sampling of micro- and nano-plastics in environmental matrixes
68. Battling the known unknowns: a synoptic review of aquatic plastics research from Australia, the United Kingdom and China
69. Chemical identification of microplastics ingested by Red Phalaropes (Phalaropus fulicarius) using Fourier Transform Infrared spectroscopy
70. Plastic ingestion by Arctic fauna: A review
71. Plastics in regurgitated Flesh-footed Shearwater (Ardenna carneipes) boluses as a monitoring tool
72. Recommended best practices for collecting, analyzing, and reporting microplastics in environmental media: Lessons learned from comprehensive monitoring of San Francisco Bay
73. Plastic debris ingestion by seabirds on the Korean Peninsula
74. Plastic ingestion by marine fish is widespread and increasing
75. Ingestion of anthropogenic materials by yellow-legged gulls (Larus michahellis) in natural, urban, and landfill sites along Portugal in relation to diet composition
76. Plastic ingestion and dispersion by vultures may produce plastic islands in natural areas
77. Review of plastic pollution policies of Arctic countries in relation to seabirds
78. Polymer types ingested by northern fulmars (Fulmarus glacialis) and southern hemisphere relatives
79. Marine pollution in fledged Leach's storm-petrels (Hydrobates leucorhous) from Baccalieu Island, Newfoundland and Labrador, Canada
80. Seasonal ingestion of anthropogenic debris in an urban population of gulls
81. Decadal changes in plastic litter regurgitated by albatrosses and giant petrels at sub-Antarctic Marion Island
82. Microplastic accumulation in the gastrointestinal tracts in birds of prey in central Florida, USA
83. Plastic ingestion by four seabird species in the Canadian Arctic: Comparisons across species and time
84. A Horizon Scan of research priorities to inform policies aimed at reducing the harm of plastic pollution to biota
85. Sampling and Quality Assurance and Quality Control: A Guide for Scientists Investigating the Occurrence of Microplastics Across Matrices
86. Quantitative overview of marine debris ingested by marine megafauna
87. Polarquest 2018 Expedition: Plastic Debris at 82°07’ North
88. Microplastic study reveals the presence of natural and synthetic fibres in the diet of King Penguins (Aptenodytes patagonicus) foraging from South Georgia
89. Plastic debris in marine birds from an island located in the South China Sea