A human impact metric for coastal ecosystems with application to seagrass beds in Atlantic Canada
Abstract
Coastal biogenic habitats are vulnerable to human impacts from both terrestrial and marine realms. Yet the broad spatial scale used in current approaches of quantifying anthropogenic stressors is not relevant to the finer scales affecting most coastal habitats. We developed a standardized human impact metric that includes five bay-scale and four local-scale (0–1 km) terrestrial and marine-based impacts to quantify the magnitude of anthropogenic impacts to coastal bays and nearshore biogenic habitats. We applied this metric to 180 seagrass beds (Zostera marina), an important biogenic habitat prioritized for marine protection, in 52 bays across Atlantic Canada. The results show that seagrass beds and coastal bays exist across a wide human impact gradient and provide insight into which are the most and least affected by human threats. Generally, land alteration, nutrient loading, and shellfish aquaculture were higher in the Gulf of St. Lawrence, whereas invasive species and fishing activities were higher along the Atlantic coast. Sixty-four percent of bays were at risk of seagrass decline from nitrogen loading. We also found high within-bay variation in impact intensity, emphasizing the necessity of quantifying impacts at multiple spatial scales. We discuss implications for management and conservation planning, and application to other coastal habitats in Canada and beyond.
Introduction
Over past decades and centuries, the magnitude, spatial extent, and variety of human impacts have substantially increased in coastal ecosystems around the world (Lotze et al. 2006; Halpern et al. 2008). Nearshore biogenic habitats, such as seagrass meadows, kelp forests, rockweed beds, and oyster reefs, are especially vulnerable as they are subject to anthropogenic threats from both the terrestrial and marine realms (Orth et al. 2006; Worm and Lotze 2006; Waycott et al. 2009; Beck et al. 2011; Krumhansl et al. 2016). Coastal management strategies have recognized nearshore biogenic habitats as areas of high conservation value, and their inclusion in marine protected areas (MPAs) is a conservation priority worldwide (DFO 2007; Cullen-Unsworth and Unsworth 2016). Despite this, it remains unclear how to prioritize areas for protection given the multitude of anthropogenic stressors impacting these ecosystems. Metrics of anthropogenic stressors used to inform management and conservation have previously been applied to ocean ecosystems across broad global and regional scales (Ban and Alder 2008; Halpern et al. 2008; Murray et al. 2015). However, these assessments are not relevant at smaller spatial scales, such as specific coastal bays, nearshore ecosystems, or biogenic habitats.
Impact metrics useful for the management and conservation planning of coastal ecosystems, particularly for specific biogenic habitats, should quantify impacts at both bay-wide and local scales. The bay-wide scale is generally defined as the geographic boundary of a bay or estuary, whereas the local scale should be relevant to the biogenic habitat in question. Human impacts are widely recognized as having scale-dependent effects on ecosystem processes (Yates and Bailey 2010; Powell et al. 2013), and spatial scale may also be relevant when assessing the magnitude of stressors influencing marine ecosystems (Thrush et al. 1999). For example, human activities were found to influence seagrass ecosystems mainly at the local scale, typically within 1–3 km of the seagrass bed (Skinner et al. 2013; Shelton et al. 2017; Cullain et al. 2018a; Iacarella et al. 2018). Yet, given that coastal biogenic habitats are often confined to bays and estuaries they are also influenced by stressors that operate at the bay scale, such as nutrient and sediment loading and pollution run-off from the surrounding watersheds (McIver et al. 2019). Additionally, various biogenic habitat types may be differentially affected by human activities, suggesting the need for habitat-specific assessments. A comprehensive assessment of human impacts in coastal ecosystems should thus consider impacts to specific habitats at both the relevant local and bay-wide scales.
In Canada, eelgrass (Zostera marina) has been designated an Ecologically Significant Species (ESS), and the inclusion of eelgrass beds within MPA networks is a central conservation priority (DFO 2009a). Given their global distribution and critical role in ecosystem functioning (Duarte 2002; Hughes et al. 2009), eelgrass beds are an ideal case study for the development of a human impact metric relevant for coastal ecosystems. Global declines in seagrass cover and associated ecosystem health have been attributed to various human activities, including nutrient pollution (Orth et al. 2006), the spread of invasive species (Williams 2007; Wong and Vercaemer 2012), coastal land alteration (Grech et al. 2012), construction of overwater structures (Fresh et al. 2006; Thom et al. 2011), and aquaculture (Skinner et al. 2013; Cullain et al. 2018a). Although natural events are also sometimes responsible for large-scale seagrass losses (i.e., ice scour and storms; Duarte 2002), human activities are recognized as a significant driver of seagrass ecosystem degradation (Short and Wyllie-Echeverria 1996; Hemminga and Duarte 2000; Waycott et al. 2009). A metric that quantifies multiple human impacts at spatial scales relevant for seagrass beds will aid conservation planning by identifying priority areas with low human impacts and highlight areas where management measures should be considered.
The objectives of this study were to (1) develop a general standardized human impact metric that can quantify the magnitude and range of anthropogenic impacts on various coastal ecosystems and biogenic habitats, and (2) apply this metric to seagrass beds in Atlantic Canada. First, we selected relevant anthropogenic impacts known to influence biogenic habitats, in particular seagrass beds, on local or bay-wide scales based on the published literature. We then compiled empirical data for these impacts for 180 seagrass beds inhabiting 52 bays across two biogeographic regions in Atlantic Canada and assessed the distribution of these impacts across the different seagrass beds, coastal bays, and biogeographic regions. Finally, we explored the potential general utility of our human impact metric for other biogenic habitats, coastal ecosystems, and geographic locations to inform coastal management and conservation.
Methods
General human impact metric development
We began by selecting relevant anthropogenic impacts for inclusion in the human impact metric for coastal ecosystems and biogenic habitats. Here, we focused on impacts known to influence seagrass beds on local or bay-wide scales and relevant indicators or measurable proxies based on the published literature (Table 1). We then assessed data availability to quantify the extent of each impact across our study region. To standardize the human impact metric across all sites, we focused on impacts and indicators for which comparable data were available from the three relevant provincial governments (Nova Scotia, New Brunswick, and Prince Edward Island) or the two marine management regions of Fisheries and Oceans Canada (Maritimes region and Gulf region), which are also considered separate biogeographic regions (Scotian Shelf and Gulf of St. Lawrence, respectively) with unique oceanographic and hydrodynamic processes (Spalding et al. 2007; DFO 2009b).
Table 1.
| Impact | Indicator | Rationale |
|---|---|---|
| Coastal land protection | Extent of protected coastal land | • Prevents erosion, reduces sediment, nutrient and pollution run-off, and maintains coastal buffer zones (Lotze et al. 2006). May be more beneficial to coastal ecosystems than marine protection (Quiros et al. 2017). |
| Watershed land use | Watershed land alteration: 1) Agriculture cover 2) Urban cover | • Predictor of water quality in receiving bays and estuaries (Uriarte et al. 2011; Huang et al. 2015). • Reliance on high water clarity makes biogenic habitats vulnerable to pollution and sediment runoff that follow conversion of natural to human dominated lands (Bryce et al. 2010; Quiros 2016; Leitão et al. 2018). |
| Human population density | Watershed human population density | • General predictor of intensity of human impacts in coastal ecosystems (Shelton et al. 2017; Iacarella et al. 2018). |
| Nutrient pollution | Human-derived nitrogen input: 1) N loading rate 2) delta-N | • Nutrients from point- and non-point sources and resulting eutrophication is a major driver of seagrass decline (Hauxwell et al. 2003; Orth et al. 2006; Waycott et al. 2009). • N-loading rate represents intensity of nutrient pollution (McIver et al. 2015), delta-N accounts for dilution from freshwater inflow and tidal exchange (Bugden et al. 2014). |
| Invasion extent | Extent of invasive biofouling species | • Non-native species increasing in coastal waters (Lotze et al. 2006). • Biofouling invaders affect biogenic habitats through shading and breakage (Brush and Nixon 2002; Wong and Vercaemer 2012). • Represents general invasion intensity as non-native species can facilitate others (Simberloff and Von Holle 1999; Braga et al. 2018). |
| Coastal commercial fishing activity | Intensity of lobster fishing | • Proxy for general fishing intensity as lobster is largest coastal fishery in Atlantic Canada (Milewski et al. 2018). • Affects biogenic habitats through direct destruction from gear, disturbance from boat traffic, and release of wastewater or chemical pollutants (Uhrin et al. 2005; Anderson et al. 2011). |
| Aquaculture activity | Presence of shellfish aquaculture facilities: 1) Near-field (<25 m) 2) Mid-field (<1 km) | • Has rapidly expanded in Atlantic Canada and range often overlaps with biogenic habitats (Bastien-Daigle et al. 2007; Comeau 2013). • Shading and physical damage occur at near-field scale (Vance 2014). Dispersed negative effects occur within 1 km radius (Skinner et al. 2013). • Far-field (bay scale) effects not considered as these include complex suite of negative and positive effects (Guyondet et al. 2013). • Finfish aquaculture accounted for in N loading estimates (McIver et al. 2018; Nagel et al. 2018). |
| Water quality | Fecal coliform counts | • Associated with reduced water clarity, decreased oxygen, and are indicative of disease-causing pathogens for coastal fauna (Parker et al. 2016; Arasamuthu et al. 2017; Lamb et al. 2017). |
| Overwater structures | Human-made overwater structures | • Structures supported above or floating on water (docks, marinas, causeways, etc.). • Causes shading, altered circulation, sediment accumulation, boating activity, and noise pollution (Fresh et al. 2006; Burgin and Hardiman 2011; Simpson et al. 2016). Suggested as the largest impact on coastal biogenic habitat health (Thom et al. 2011; Rehr et al. 2014). |
| Riparian land alteration | Human altered riparian land | • Defined as land within 0–50 m from shoreline (Quiros et al. 2017). • Causes sediment instability, fine sediment loading, and negative consequences for biogenic habitats and associated fauna (Bryce et al. 2010; Van Katwijk et al. 2011; Quiros 2016; Leitão et al. 2018). |
We separated impacts into those acting at a bay scale or local scale. The bay scale was defined by the geographic boundary of the bay (or estuary) and considered impacts that influence the entire waterbody of the bay, for example through tidal mixing or indirect effects, thereby also influencing the biogenic habitats within. In our case, this included watershed land use, human population density, nutrient pollution, invasion extent, and coastal commercial fishing activity (Fig. 1). In contrast, local-scale impacts were defined as those within a local zone of influence. Previous studies have found a local scale of <1 to <3 km as being most relevant for assessing human impacts to seagrass beds (Skinner et al. 2013; Shelton et al. 2017; Iacarella et al. 2018). We included shellfish aquaculture activity, water quality, overwater structures, and riparian land alteration (Fig. 1) in close vicinity to our seagrass beds as local-scale impacts. These impacts were all measured within a 0–1 km distance of the seagrass beds as a conservative estimate and to distinguish from broader bay-scale impacts.
Figure 1.
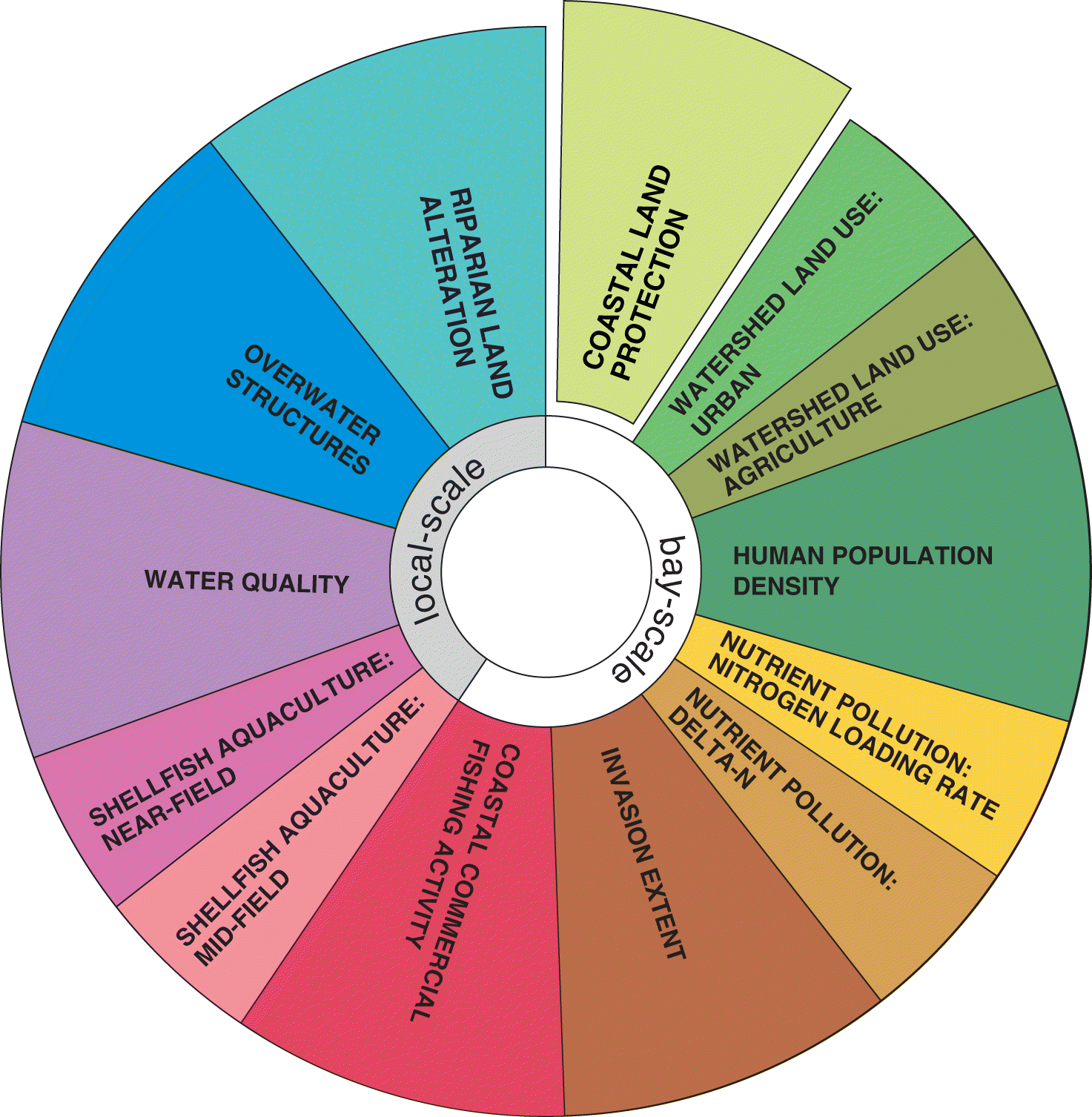
Our derived human impact metric therefore included five bay-scale and four local-scale impacts, with three of these impacts (land use, nutrient pollution, and aquaculture activity) each further subdivided, resulting in a total of 12 impact scores (Fig. 1). In addition, we included coastal land protection as a measure expected to benefit our biogenic habitat. Depending on the coastal ecosystem or biogenic habitat in question, these impacts and indicators and the scale at which they are assessed can be easily adapted. See Table 1 for the detailed rationale for the inclusion of these impacts in our human impact metric for seagrass beds in Atlantic Canada. In the discussion, we further describe the potential inclusion of other relevant human impacts once quantitative data becomes publicly available.
Data collection
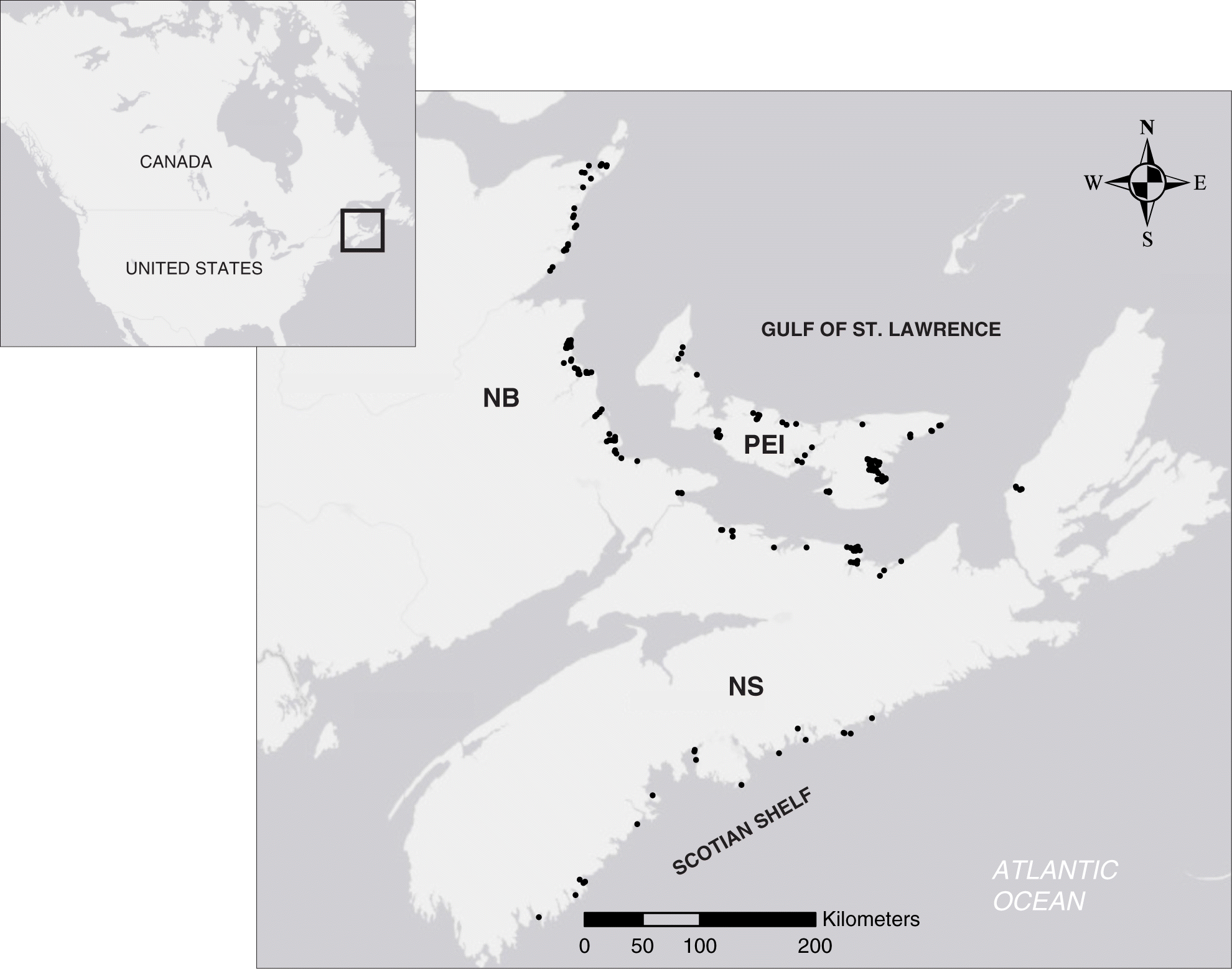
Our goal was to apply the human impact metric to seagrass beds across Atlantic Canada. To do so, we first compiled the locations of 180 seagrass beds (Fig. 2) from field surveys conducted over the past decade (Weldon et al. 2009; Schmidt et al. 2012; Skinner et al. 2013; Cullain et al. 2018b; Wong 2018; A. Locke and R. Bernier, personal communication, 2018) in 52 bays along the coasts of Nova Scotia (NS), New Brunswick (NB), and Prince Edward Island (PEI) and in two biogeographic regions (Scotian Shelf, Gulf of St. Lawrence).
Figure 2.
Coastal land protection
We obtained GIS shapefiles of provincial, federal, and private conservation areas from Environment and Climate Change Canada. We clipped shapefiles to the 0–200 m coastal land surrounding each bay to determine the percentage of protected land within each bay’s coastal land zone. We defined coastal land as 0–200 m from the coastline because this distance has previously been identified as important for pollution input to receiving waters (Valiela et al. 1997).
Land use
We used previously delineated watershed boundaries for each bay provided by provincial authorities (NS Department of Environment, NB Department of Energy and Resource Development, PEI Department of Land and Communities). In some cases, we further delineated watersheds to include all freshwater inputs by predicting bay-specific watercourse drainage patterns using hydrographic data and digital terrain models (GeoNova, GeoNB, PEI GIS Data Catalog). We clipped shapefiles that classified land use types across each of the provinces to the predetermined watershed areas for each bay. The percentages of urban (including industrial and commercial use) and agriculture land use were determined by summing appropriate units in the watershed areas and dividing by total watershed area. We also determined the number of bays with >10% combined urban and agriculture watershed land use as this threshold has been identified as presenting a moderate to high risk for ecosystem degradation in receiving waters (WWF 2017) and has been used to differentiate developed vs. undeveloped watershed land (Lerberg et al. 2000; Bilkovic et al. 2006; Blake et al. 2014).
Human population density
Watershed human population density was estimated using the number of civic addresses present in each watershed (GeoNova, GeoNB, PEI GIS Data Catalog) multiplied by the average number of residents per household (2.3, 2.3, and 2.4 in NS, NB, and PEI, respectively) (Statistics Canada 2017). The number of individuals was standardized to watershed area.
Nutrient pollution
Nutrient pollution was divided into two sub-measures: nitrogen loading rate and delta-N (ΔN). We used data from previous applications of a nitrogen loading model (NLM), originally developed for Waquoit Bay, Massachusetts (Valiela et al. 1997), for several bays in Atlantic Canada, including seven bays in NB (McIver et al. 2015), 21 bays in NS (Nagel et al. 2018), and six bays in PEI (Palmer 2018). For the remaining 19 bays, we estimated total nitrogen load (kg N year−1) using linear regression models specific to each province (Supplementary Material 1, Figs. S1 and S2). Briefly, the NLM estimates watershed and atmospheric derived total dissolved nitrogen loads from several point (i.e., direct atmospheric deposition, wastewater discharge, seafood processing plants, finfish aquaculture) and non-point sources (i.e., indirect atmospheric deposition, septic systems, and fertilizer addition) with appropriate loss parameters (Valiela et al. 1997; McIver et al. 2015, 2018). Final nitrogen loading rates were calculated by summing across all sources and standardized to the area of each bay (kg N ha bay−1 year−1) to account for dilution effects. We assessed how many bays had nitrogen loading rates above the 50 kg N ha bay−1 year−1 threshold identified by Latimer and Rego (2010) as being detrimental to seagrass coverage.
The ΔN combines total nitrogen load (kg N year−1, as calculated above) with a bay’s tidal flushing time and freshwater recharge volume to estimate the increase in nitrogen concentration above ambient oceanic nitrogen concentration after dilution factors (Bugden et al. 2014). We calculated tidal flushing and freshwater recharge for each bay as described in Nagel et al. (2018). We assessed how many bays had ΔN values above the 0.06 threshold identified by Bugden et al. (2014) as being likely to experience anoxic events as a result of excess nitrogen input.
Invasion extent
We used presence/absence data from the Aquatic Invasive Species (AIS) monitoring program (Sephton et al. 2017) to estimate invasion extent for each bay. This program monitors the presence of nine invasive biofouling species by assessing settlement on plastic monitoring plates. These species included seven tunicates: vase tunicate (Ciona intestinalis), clubbed tunicate (Styela clava), carpet tunicate (Didemnum vexillum), golden star tunicate (Botryllus schlosseri), violet tunicate (Botrylloides violaceous), compound sea squirt (Diplosoma listerianum), and European sea squirt (Ascidiella aspersa), a bryozoan (Membranipora membranacea) and an amphipod, the Japanese skeleton shrimp (Caprella mutica). Although the European green crab (Carcinus maenas) is also invasive in our study region and can negatively impact seagrass beds (Garbary et al. 2014), we were unable to obtain quantitative estimates of green crab densities for each of our sites to include in our quantification of invasion extent, but note that green crabs are known to occur in all 52 bays.
Our metric of invasion extent is a measure of invasive biofouling species richness over 10 years. We used a 10-year average (2006–2015) of invader presence/absence for each of the nine invasive biofouling species and averaged across species to obtain a measure of invasion extent for each bay. When AIS monitoring stations were not present in a bay we used the closest station within 50 km. When no monitoring station was located within 50 km for a specific year we excluded that year from the 10-year average. As the AIS monitoring stations are not located within seagrass beds, our measure of invasion extent is a proxy for how likely fouling is expected within a bay and is not indicative of invaders specifically fouling seagrass at that site (Table 1).
Coastal commercial fishing activity
We used data from Fisheries and Oceans Canada to calculate the number of inshore and offshore lobster fishing licenses per port in 2018 as an estimate of lobster fishing intensity (A. Cook, personal communication, 2018). Although these data do not indicate where lobster fishing is taking place (i.e., inshore, offshore, or in an adjacent bay), they provide a general estimate of the extent of fishing vessel traffic.
Aquaculture activity
Shellfish aquaculture activity was measured as presence/absence. We used data from the NS Department of Fisheries and Aquaculture; the NB Department of Agriculture, Aquaculture, and Fisheries; and Fisheries and Oceans Canada to determine the presence or absence of operational shellfish aquaculture leases in 2018 within a <25 m radius (near-field) and <1 km radius (mid-field) of each seagrass bed. The impact of finfish aquaculture was included in nutrient loading estimates as outlined above.
Water quality
We used fecal coliform monitoring data from the Canadian Shellfish Sanitation Program (CSSP 2016) as a measure of water quality. We used a 10-year average (2005–2015) of fecal coliform counts (Most Probable Number (MPN) 100 mL−1) from the CSSP monitoring station closest to each seagrass bed, which were typically <500 m away. We assessed water quality contamination according to thresholds set by CCSP as fecal coliform counts <14 MPN 100 mL−1 considered uncontaminated and of good quality (CSSP 2016).
Overwater structures
We manually classified the total area of overwater structures (i.e., wharfs, bridges, causeways, etc.) within a 1 km radius of each seagrass bed using Google Earth (Google Inc. 2018). We standardized the total area of overwater structures to the water surface area within a 1 km radius.
Riparian land alteration
Using the land use data described above, we estimated human altered riparian land within 0–50 m from the coastline due to urban, commercial, industrial, agriculture, and forestry land use in contrast with unaltered natural wetland, forest, or conservation land areas. We define riparian land as 0–50 m from the coastline because this distance has previously been used to define riparian land and human land alteration within this range has been shown to be detrimental to seagrass health (Quiros et al. 2017). We measured the area of riparian land alteration within a 1 km radius of seagrass, which was standardized to the total riparian land area within the 1 km radius of each seagrass site. We used the same land alteration thresholds for watershed land use (>10%) to assess how many seagrass beds had adjacent riparian land considered a moderate to high risk for ecosystem degradation in receiving waters (WWF 2017). This threshold is based on total watershed land alteration, as opposed to strictly riparian land, as no threshold estimates exist for riparian land alteration. Given that coastal land alteration near seagrass beds is expected to be more detrimental than alteration in the entire watershed (Quiros et al. 2017), we expect this to be a conservative threshold.
Human impact standardization
We used the above data to first calculate the intensity of each human impact at each of the 180 seagrass beds (raw data). Each impact was then standardized to range from 0 to 1, and the multiple impacts at each site were compiled into a petal diagram to illustrate the overall human impact level. Standardization was performed at two different spatial scales: across all 52 bays or 180 sites and across each region.
We used R version 3.2.2 (R Core Team 2015) and the ggplot2 package (Wickham 2016) to assess the distribution of raw impact scores across all bays and seagrass beds. We determined significant differences between the median scores for regions based on 95% confidence intervals, which are displayed in notched boxplots. Petal diagrams were created in Microsoft Excel and CorelDraw version 19.
Results
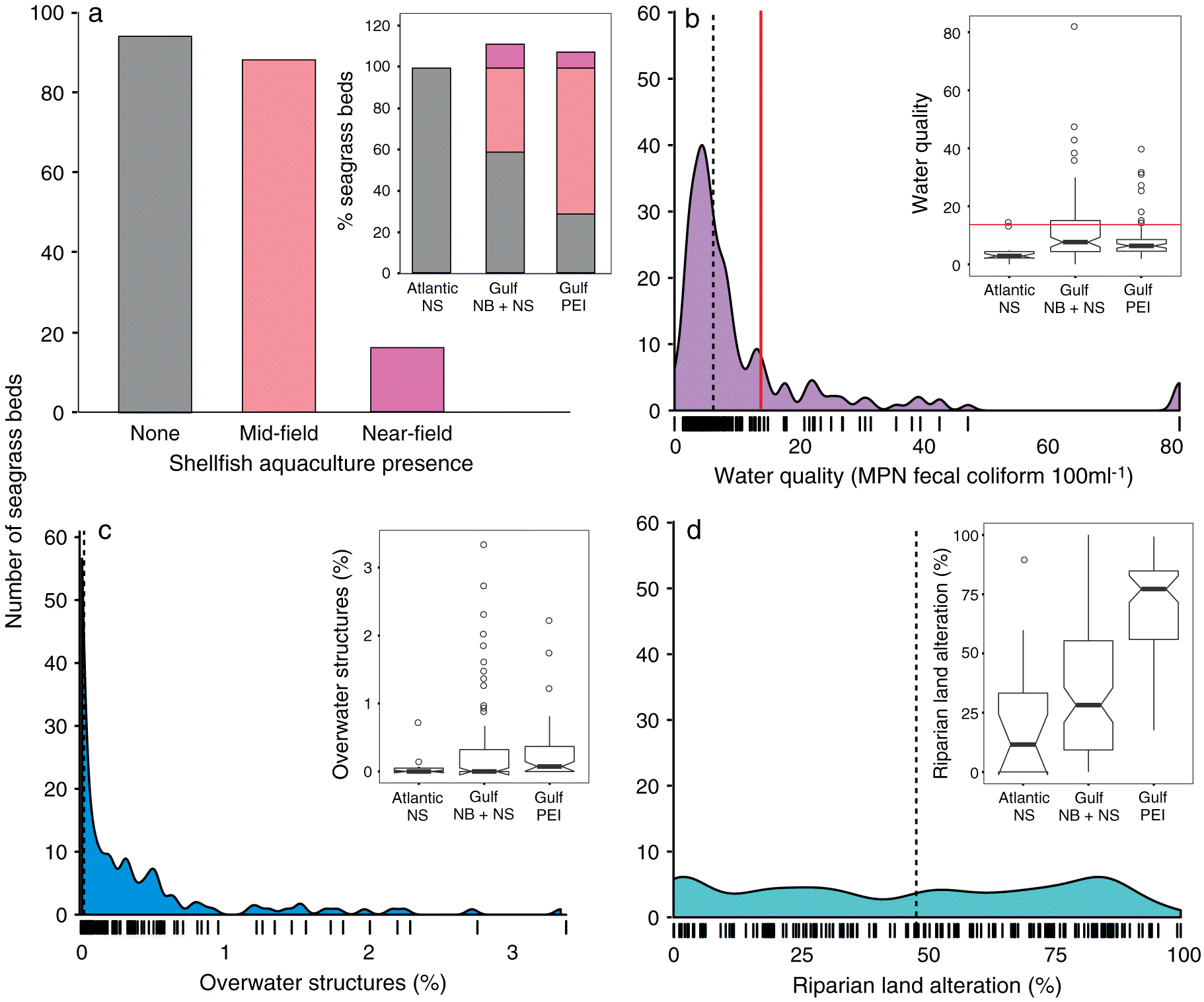
Comparing the intensity of human impacts across all 52 coastal bays (Fig. 3) and 180 seagrass beds (Fig. 4) revealed a wide gradient of human impacts in Atlantic Canada. We observed differences in the distribution of each impact between the Scotian Shelf and Gulf of St. Lawrence, and within the Gulf region between the mainland coast (NB and NS) and PEI (Figs. 3 and 4 insets). Below we describe and assess these differences among regions (Atlantic NS, Gulf NB + NS, and PEI) based on 95% confidence intervals (Figs. 3 and 4 insets).
Figure 3.
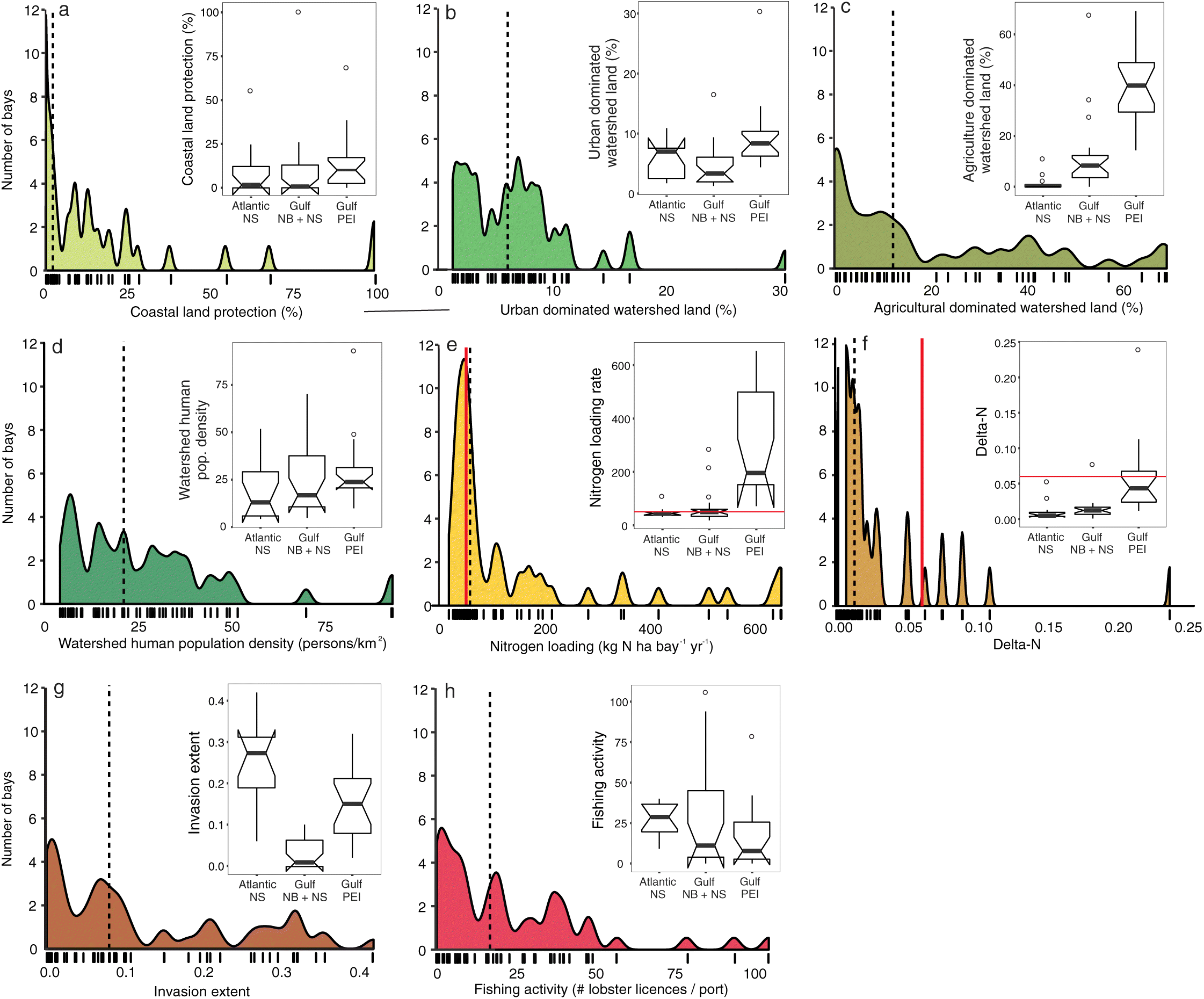
Figure 4.
Bay-scale impacts
Land protection and use
The percentage of coastal land protection was highly variable, ranging from 0% to 100% protection (median = 3.21%; Fig. 3(a)). Coastal land protection was highest in PEI (10.04% ± 5.42% 95% CI), much greater than in the Gulf NB + NS (0.68% ± 4.32%) and Atlantic NS (1.65% ± 5.53%), although not statistically significant. PEI watersheds also had the greatest land alteration for urban (Fig. 3(b)) and agricultural uses (Fig. 3(c)), although urban land use was similar between PEI and Atlantic NS (8.3 ± 1.49% and 7.1% ± 2.23%, respectively), but significantly greater than the Gulf NB + NS (3.5% ± 1.35%). Agricultural land use in PEI (39.8% ± 7.11%) was significantly greater than in the Gulf NB + NS (8.4% ± 2.87%) and Atlantic NS (3.3% ± 0.32%). According to the threshold of >10% watershed land alteration (WWF 2017), 65% of the 52 bays were at risk for ecosystem degradation in receiving waters, most of them in the Gulf NB + NS and PEI, and only two in Atlantic NS.
Human population density
Similar to the regional variation in land protection and use, the median human population density was higher in PEI (24.1 persons km−2) than in Atlantic NS (13.2 km−2) and the Gulf NB + NS (16.9 km−2), although not statistically different among regions (Fig. 3(d)).
Nutrient pollution
The median human derived nitrogen loading rate across the 52 bays was 58.9 ± 25.9 kg N ha bay−1 year−1. Nitrogen loading rates were significantly higher in PEI bays (193.7 ± 122.22) than in Atlantic NS (45 ± 5.84) and the Gulf NB + NS (52.5 ± 8.83; Fig. 3(e)). Across all bays, 64% were above the 50 kg N ha bay−1 year−1 threshold identified by Latimer and Rego (2010) as being detrimental to seagrass coverage (Fig. 3(e)). This included all (100%) of bays in PEI, 55% in the Gulf NB + NS, but only 25% in Atlantic NS. A similar pattern was observed for ΔN, indicating that PEI bays had the highest eutrophication risk (0.041 ± 0.016; Fig. 3(f)), followed by the Gulf NB + NS bays (0.012 ± 0.003), although not significantly different, whereas ΔN in Atlantic NS bays (0.005 ± 0.003) was significantly lower than in both PEI and the Gulf NB + NS. Only 15% of all bays had ΔN values above the 0.06 threshold that would indicate high probability of anoxic events (Bugden et al. 2014). Again, most bays above this threshold were located in PEI, whereas only one Gulf NB + NS bay (River Philip) and no Atlantic NS bays were considered at risk for anoxic events.
Invasion extent
The invasion extent of non-native biofouling species was significantly higher in Atlantic NS bays (median = 0.28 ± 0.05; Fig. 3(g)) than in the Gulf bays. The invasion extent in PEI (0.15 ± 0.05) was also significantly higher than in Gulf NB + NS (0.01 ± 0.02). The dominant invaders across the three regions were vase tunicates, violet tunicates, golden star tunicates, and Membranipora membranacea. However, M. membranacea was not detected in any PEI bays, and European sea squirts were only detected in Atlantic NS bays.
Coastal commercial fishing activity
Lobster fishing was significantly higher in Atlantic NS bays (median = 29 ± 7.6 licenses port−1) than in PEI (8 ± 8.4), whereas that in the Gulf NB + NS (11 ± 13.5) was not significantly different from the other regions (Fig. 3(h)).
Local-scale impacts
Aquaculture activity
Near-field shellfish aquaculture (<25 m from the seagrass bed) was present at 20 seagrass beds, and mid-field (<1 km) at 87 beds across all 180 sites (Fig. 4(a)). Among regions, Atlantic NS had no shellfish aquaculture within 1 km of any seagrass beds, and mid-field aquaculture was more prevalent in PEI (71% of beds) and the Gulf NB + NS (41%) than near-field (8% and 15%, respectively).
Water quality
Across all 180 sites, fecal coliform counts close to seagrass beds ranged from 0 to 82 MPN 100 mL−1 (median = 6.1 ± 1 MPN 100 mL−1, Fig. 4(b)). Median fecal coliform counts near seagrass beds were significantly lower in Atlantic NS (2.58 ± 0.8 MPN 100 mL−1) than in PEI (6.1 ± 0.8) and the Gulf NB + NS (7.62 ± 1.7; Fig. 4(b)), indicating that, overall, water quality is uncontaminated and of good quality according to the CCSP threshold (<14 MPN 100 mL−1). However, 35 (19%) of seagrass beds had water quality above this threshold, with one in Atlantic NS, nine in PEI, and 25 in the Gulf NB + NS.
Overwater structures
The percentage of water covered by overwater structures near seagrass beds ranged from 0% to 3.38% across all seagrass beds (Fig. 4(c)). The median percentage was greater in PEI (0.074% ± 0.07%) than in the Gulf NB + NS (0.00% ± 0.03%) and Atlantic NS (0.00% ± 0.02%), with no statistical differences among regions (Fig. 4(c)).
Riparian land alteration
Similar to human land alteration in the entire watershed, the median percentage of coastal land altered in close proximity to seagrass beds was significantly higher in PEI (7.72% ± 5.6%, Fig. 4(d)) than in Atlantic NS (11.5% ± 12.8%) and the Gulf NB + NS (28.1% ± 7.3%). However, the range of coastal land alteration was similar in all three regions. Of the 180 seagrass beds, 82% were affected by >10% riparian land alteration.
Human impacts at different spatial scales
Our human impact metric can be compared across all coastal bays and seagrass beds in Atlantic Canada (Figs. 3 and 4) as well as within biogeographic regions (e.g., the Scotian Shelf and Gulf of St. Lawrence, Figs. 5 and 6). Standardizing the impacts within each region allows for better comparison of the importance of each impact relative to other sites subjected to similar biogeographic conditions. For example, compared with all other seagrass beds, those on the Atlantic coast of NS had relatively low human impact (Figs. 3 and 4 insets); yet when standardized within the Scotian Shelf region, relative impact patterns became more apparent (Fig. 5). This revealed that several seagrass beds were minimally impacted by human activities relative to other Atlantic NS sites, including Port Joli, Cable Island, and Taylor’s Head, which have higher coastal protection and lower overall bay and local-scale impacts (Fig. 5). In comparison, other seagrass beds were more impacted by human activities, with Second Peninsula highly impacted by urban and agriculture land use, poor water quality, and commercial fishing; St. Margaret’s Bay by riparian land alteration, poor water quality, and commercial fishing; Sambro by high population density and urban land use; Port Mouton by commercial fishing and overwater structures; and Musquodoboit by nutrient pollution (Fig. 5).
Figure 5.
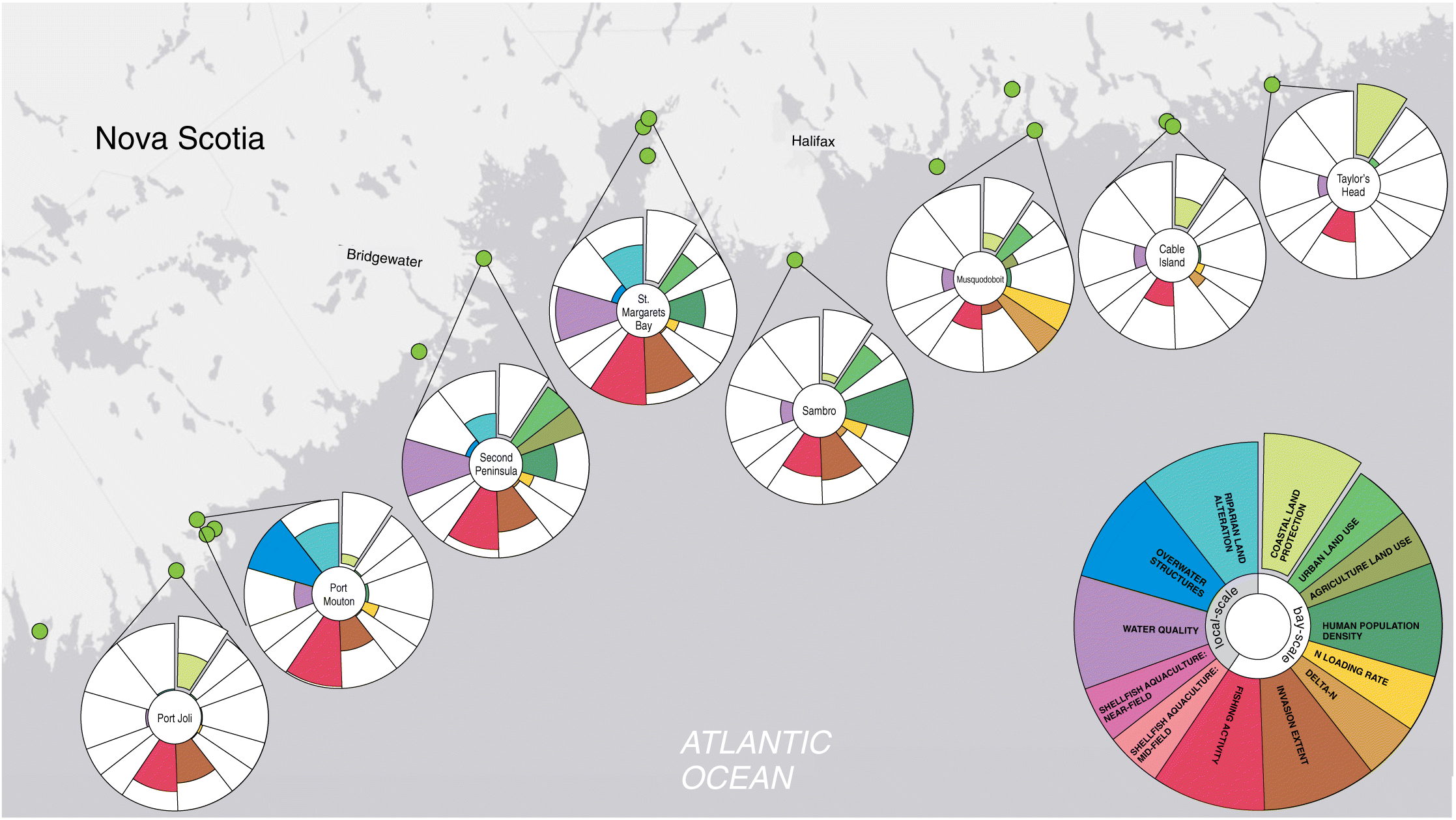
Figure 6.
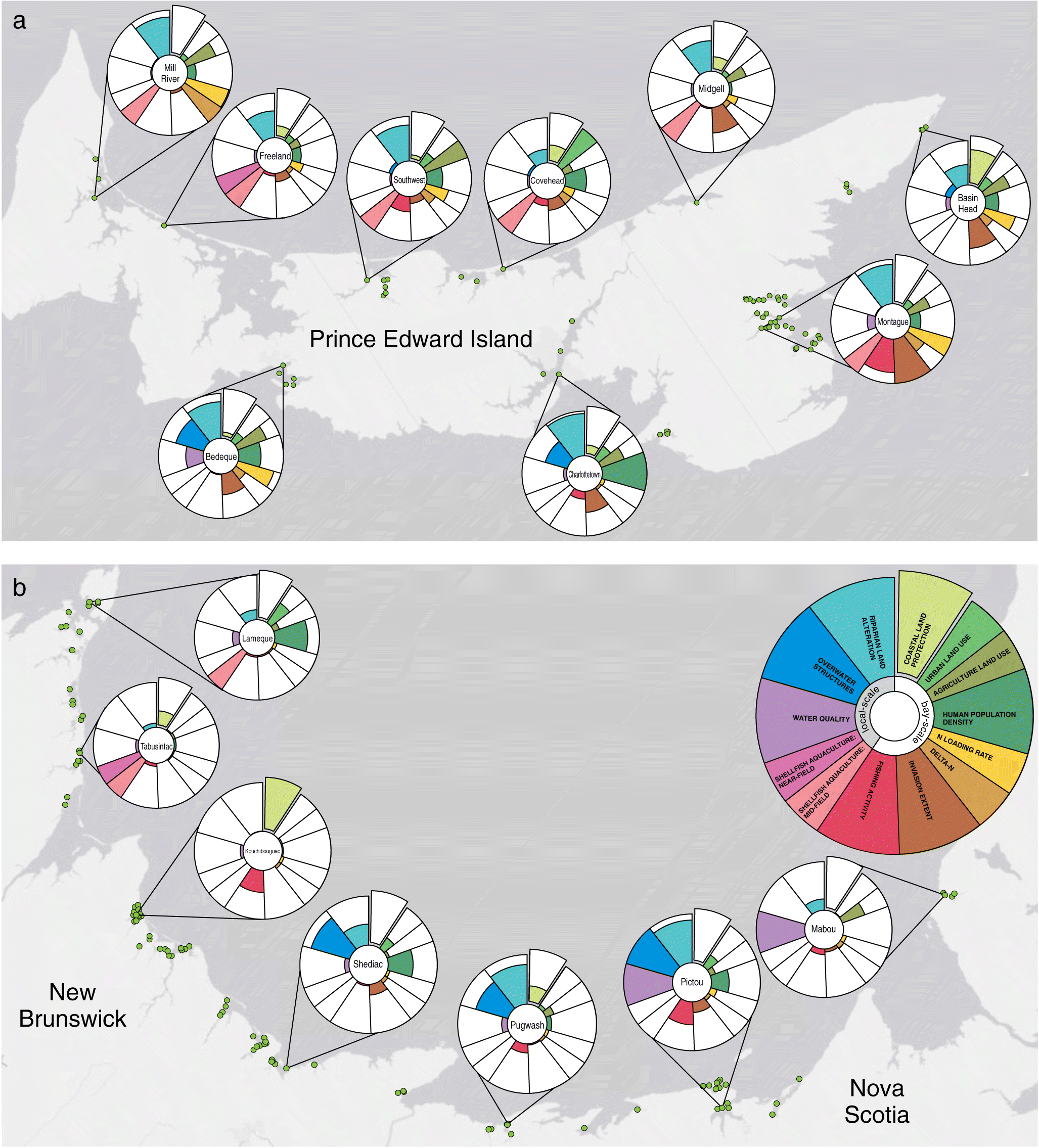
When standardized within the Gulf region the impact metric also revealed many seagrass beds highly impacted by human activities (Fig. 6). For example, seagrass beds in Pictou were highly impacted by poor water quality and overwater structures, Mill River had high riparian land alteration, and Charlottetown had high human population density (Fig. 6). In contrast, other seagrass beds were minimally impacted by human activities, including those in Kouchibouguac National Park; however there were fewer minimally impacted beds in the Gulf region (Fig. 6) compared with the Scotian Shelf region (Fig. 5).
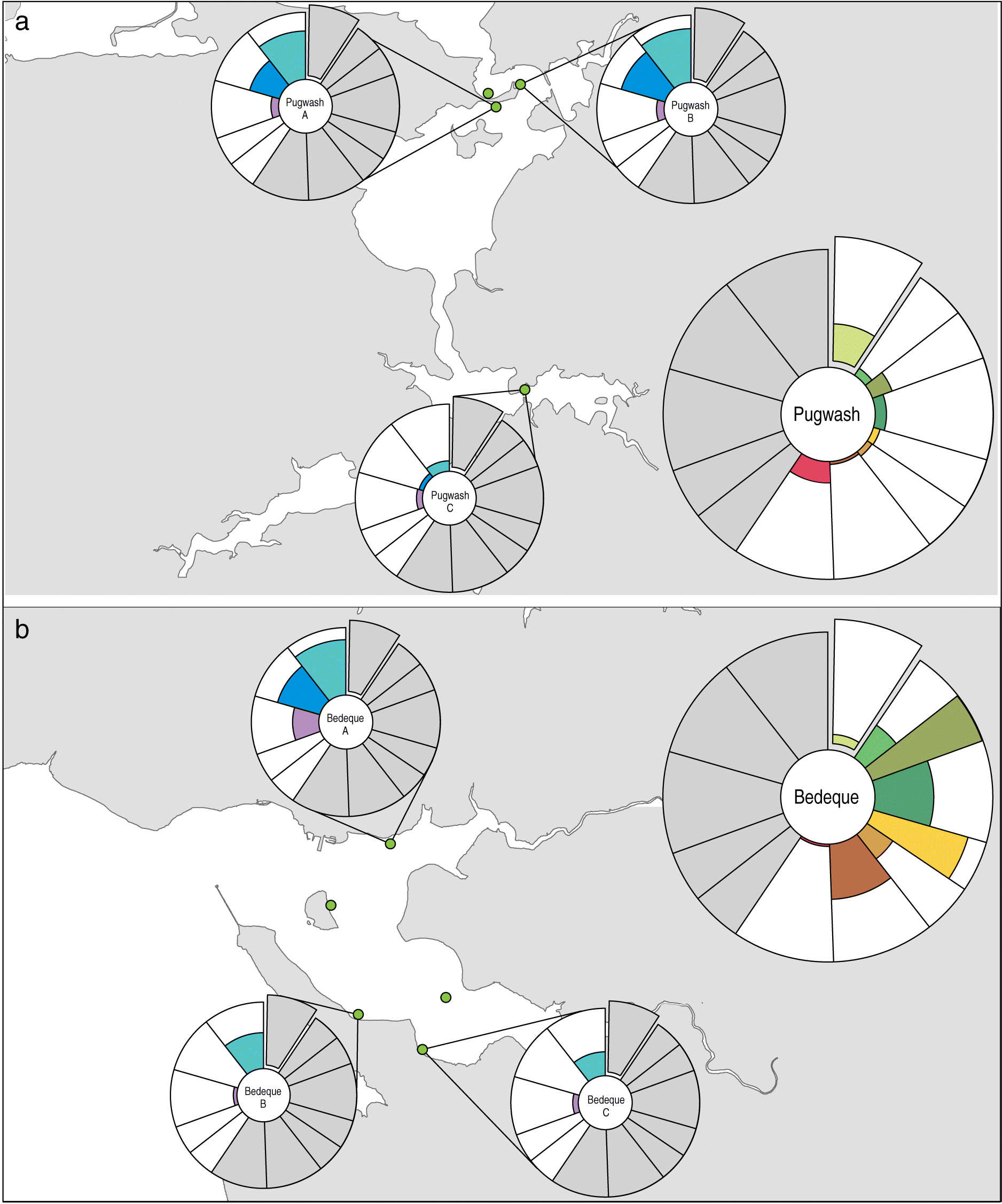
Our human impact metric can also be used to assess the degree of impacts among individual seagrass beds within a bay. For example, Pugwash Bay (Fig. 7(a)) had relatively low human impacts at the bay-scale (i.e., almost all impacts within the 20th–40th percentile), but high heterogeneity in local-scale impacts among the individual seagrass beds. For example, Bed “C” (Fig. 7(a)) was only minimally impacted by poor water quality and overwater structures, whereas beds “A” and “B” were more highly impacted by overwater structures and reduced buffer zones. In comparison, Bedeque Bay in PEI (Fig. 7(b)) was highly impacted by human activities at the bay scale (i.e., most bay-scale impacts in the 80th–98th percentile) with high agricultural land use and nitrogen loading. Again, high heterogeneity in human impacts was evident among the individual seagrass beds (Fig. 7(b)). For example, Bed “A” had extensive overwater structures in its vicinity and poor water quality, whereas beds “B” and “C” were influenced by a relatively low degree of human impacts at the local scale.
Figure 7.
Discussion
Quantifying the magnitude and range of human impacts in a standardized, comprehensive and comparable way is urgently needed for large-scale scientific assessments and for management and conservation planning in coastal ecosystems. Previous metrics of human impacts for marine ecosystems have typically been developed on global, national, or broad regional scales, with little effort towards metrics appropriate for coastal ecosystems or specific biogenic habitats. Our human impact metric incorporates a suite of anthropogenic stressors from both terrestrial and marine sources relevant for coastal bays and estuaries as well as biogenic habitats and can be adapted to and standardized across multiple spatial scales.
Despite including multiple human impacts into our metric, we did not to combine them into one cumulative impact score (sensu Halpern et al. 2008), as this requires scientifically-informed vulnerability weightings that are not readily available. Although research on human impacts to seagrass ecosystems in Atlantic Canada has increased over the past decade (Coll et al. 2011; Schmidt et al. 2012; Hitchcock et al. 2017; Cullain et al. 2018a; McIver et al. 2019), we still know too little about the responses of seagrass ecosystems to individual and cumulative human impacts to allow effective ranking or evidence-based weighting of their importance. Despite not explicitly accounting for the magnitude and relative importance of all impacts combined, our approach still provides a useful overall qualitative summary of the total human impact as visualized by the petal diagrams. Furthermore, providing individual impact scores (Supplementary Materials 2, 3, and 4) instead of collapsing these into a single value allows influential impacts to be identified for specific bays, the relative impact to be compared, and also allows managers to identify and track impacts of interest. In the following, we discuss our application of the human impact metric to seagrass beds in Atlantic Canada, and then its potential for more general application to other habitats and regions and uses for management and conservation planning.
Application to seagrass beds in Atlantic Canada
Applying our human impact metric to 180 seagrass beds in 52 coastal bays in Atlantic Canada revealed several insights relevant for scientific assessment, management, and conservation. We found considerable regional variation in impact strength that provides insight into where coastal bays and seagrass ecosystems are expected to be most and least affected by individual or cumulative human impacts. Also, our results highlight the importance of considering human impacts at multiple spatial scales when assessing threats to coastal ecosystems and biogenic habitats.
One of the most damaging human impacts to the extent and condition of seagrass beds is eutrophication from human-derived nitrogen loading (Orth et al. 2006; Waycott et al. 2009). We found that 64% of all bays in our study are at risk of seagrass decline based on nitrogen loading rates (Latimer and Rego 2010). However, only 13% of bays, most of which are in PEI, are at risk for anoxic events based on ΔN values (Bugden et al. 2014). Although both nitrogen loading and ΔN can indicate eutrophication risk, ΔN can indicate the potential for more severe anoxic events to occur by accounting for dilution from freshwater input and tidal flushing (Bugden et al. 2014). Thus, although human-derived nitrogen loading can be high in many bays and lead to primary (e.g., enhanced phytoplankton, epiphytic, or benthic algal growth) or secondary (e.g., reduced shoot density and biomass) eutrophication effects in seagrass beds (Schmidt et al. 2012; Cullain et al. 2018a; McIver et al. 2019), more severe loss of seagrass cover from anoxic events may only be of concern in PEI (Bugden et al. 2014).
Land alteration resulting from urban or agricultural land use was generally much higher in the Gulf region, particularly in PEI, and is related to higher human population density, higher nutrient loading, poorer water quality, and other human impacts. Yet, despite relatively lower land use changes within Atlantic NS watersheds, the wide range of riparian land alteration close to seagrass beds suggests that local-scale impacts may still be important for seagrass health in Atlantic NS. Additionally, 42% of Atlantic NS bays have no coastal land protection. This is similar to bays in the Gulf NB + NS but is much higher than PEI where only 11% of bays have no coastal land protection. Seagrass beds bordered by land managed for conservation purposes usually have higher temporal stability than beds associated with unprotected land (Breininger et al. 2016). Not surprisingly, our results show that coastal land protection at the bay scale is positively related to the extent of riparian land alteration adjacent to seagrass beds, i.e., local scale (p < 0.001, R2 = 0.24, Fig. S3). This suggests that the implementation of coastal conservation areas at the bay scale may be a useful strategy to reduce the number of seagrass beds at risk of reduced buffer zones from riparian land alteration (Quiros et al. 2017).
Compared with the Gulf region, Atlantic NS bays have a higher extent of invasive biofouling species. This is not surprising given that the closer proximity of Atlantic NS to the open Atlantic Ocean and United States allows greater susceptibility to range expansions of invaders (Sephton et al. 2017). The difference in invasion extent between the two regions is driven by the recent invasion of several tunicate species in NS that are still absent from the Gulf of St. Lawrence (Moore et al. 2014; Vercaemer et al. 2015). However, species distribution models predict northward range expansion of most invasive tunicates, with many seagrass-inhabited bays in the Gulf region becoming hotspots for invasive biofouling species over the next 50 years (Lowen and DiBacco 2017). Fouling by invasive tunicates reduces seagrass growth and survival because of shading and premature breakage (Brush and Nixon 2002; Wong and Vercaemer 2012). As our human impact metric identified many highly impacted seagrass beds in the Gulf region, investigating how seagrass beds respond to the cumulative effect of invasive biofouling species and other human impacts should be a research priority as this may become a reality in the near future.
Our bays along the Atlantic coast of NS also have greater lobster fishing activity compared with bays in the Gulf region. Although lobster fishing does not have direct impacts on seagrass bed health, the lobster fishery is currently the most valuable commercial fishery in Atlantic Canada with the largest fleet of small fishing boats operating in coastal waters (Milewski et al. 2018). It may, therefore, reflect the intensity of boating activity in the region, some of which can directly impact seagrass beds through physical disturbance (Uhrin et al. 2005; Anderson et al. 2011). In contrast, currently none of our seagrass beds in Atlantic NS are influenced by mid- or near-field shellfish aquaculture activity, whereas over half of seagrass beds in the Gulf region are in close proximity to shellfish aquaculture operations. Although shellfish aquaculture does occur in NS, it is not as widespread as in NB or PEI (Bastien-Daigle et al. 2007; Comeau 2013). We did not include finfish aquaculture activity as a separate impact in our metric as only two bays (Port Mouton and False Passage/Cable Island) have finfish aquaculture leases within their boundaries. Instead, the nitrogen released from finfish aquaculture, which primarily derives from fish excretory products and the breakdown of particulate organic waste, is accounted for in the nutrient pollution impact. Excess nitrogen input has been apparent in Port Mouton Bay where finfish aquaculture, when in operation, was estimated to increase anthropogenic nitrogen loading by 14% (McIver et al. 2018; Nagel et al. 2018). Therefore, the estimated nitrogen loading rate in Port Mouton over the past 10 years was higher than nearby bays (e.g., Port Joli) despite similar watershed land use and human population density and is nearing the threshold considered detrimental to seagrass (Fig. 5).
Overwater structures are recognized as having one of the largest direct negative impacts on seagrasses, yet no threshold level has been determined (Fresh et al. 2006; Rehr et al. 2014). Therefore, it remains unclear what degree of overwater structure coverage is detrimental to seagrass and if the range of impact scores we observed in Atlantic Canada are considered harmful. Although the median percentage of water covered by overwater structures was relatively low for most seagrass beds, with no structures within 1 km of 47% of seagrass beds, our impact metric identified several beds where coverage by overwater structures was much higher. These were primarily in the Gulf region and, in some cases, in bays where other human impacts were low. For example, overwater structures near some seagrass beds in Pugwash Bay, Shediac Bay, and Port Mouton are relatively high despite low impact scores for most other threats (Figs. 5 and 6(b)). This illustrates the importance of quantifying numerous threats at multiple scales to fully understand impacts to biogenic habitats. Future research that identifies thresholds for the effects of overwater structures on coastal biogenic habitats would improve assessments of ecosystem health.
Assessment of human impact at multiple spatial scales
Our human impact metric is unique as it assesses impacts at multiple spatial scales. This allows evaluation of within-bay variation and can reveal discrepancies about the relative magnitude of impacts at different spatial scales. A metric that accounts for different spatial scales is critical for biogenic habitats in coastal ecosystems given the uncertainty in what human impacts, and at what spatial scales, are most detrimental (Coll et al. 2011; Skinner et al. 2013; Shelton et al. 2017; Iacarella et al. 2018). Seagrass bed responses to anthropogenic stressors typically exhibit a high level of variability within bays (Hitchcock et al. 2017) and across regions (Coll et al. 2011; Cullain et al. 2018b; McIver et al. 2019), which is reflected in our results (Figs. 5–7). By considering impacts at both local and bay-wide scales our metric can highlight locations where there is spatial mismatch between the intensity of bay-wide and local-scale impacts at individual seagrass beds. For example, Bedeque Bay in PEI is a highly impacted bay based on bay-scale impacts; however, there is high variability among local-scale impacts within the bay (Fig. 7(b)). Observation of only local-scale impacts would suggest certain beds are relatively non-impacted when this might not be the case. This emphasizes the necessity of quantifying impacts at multiple scales to fully understand the degree of impact to coastal biogenic habitats.
General application to other habitats and ecosystems
Along with seagrass, several other species create complex biogenic habitats in coastal waters in Canada and elsewhere, including rockweed beds, kelp forests, oyster reefs, and others (Beazley et al. 2017). Despite broad ideas of how these biogenic habitats respond to human pressures, little is known of the trajectories for individual habitats, what management actions are needed, and where to prioritize conservation efforts (DFO 2018). When developing our human impact metric, we focused on impacts that are not only relevant for biogenic habitats but where similar data to quantify the extent of each impact is obtainable for other provinces and coasts in Canada, as well as other countries. For example, the data used to calculate water quality, coastal fishing activity, and invasion extent are all from federal programs that collect comparable data for other coastal regions of Canada. The model used to estimate nitrogen loading rates can be adapted to coastal bays and estuaries worldwide (Valiela et al. 1997), and the data used to calculate coastal land protection, land uses, human population density, and aquaculture presence are all provincially regulated with comparable data available across provinces. Thus, our human impact metric can be readily applied to other coastal regions and habitats in Canada and beyond. This opens the door for direct comparisons of human impacts to a specific habitat, i.e., seagrass beds, across provinces, biogeographic regions, and coasts, as well as comparisons between different biogenic habitats within and across countries.
The raw and standardized impact scores for all 180 seagrass beds and 52 bays assessed in this study (Supplementary Materials 2, 3, and 4) are available to aid in the development of monitoring programs, management strategies and prioritization of coastal conservation efforts in Atlantic Canada. This database will be valuable for the application of our human impact metric to other biogenic coastal ecosystems in Atlantic Canada, as many of the seagrass-inhabited bays are also home to kelp and rockweed habitats. Therefore, the same bay-scale impacts already calculated for the current database apply and only local-scale impacts need to be adjusted.
Our impact metric has been specifically tailored for impacts relevant to seagrass beds in Atlantic Canada. Applications to other ecosystems and regions should begin with an assessment of the impacts present and their potential influence (e.g., Table 1). For example, boat moorings are not generally a critical threat to most Atlantic Canada seagrass beds yet should be included in impact metrics for seagrass in parts of the coastal United States, United Kingdom, and Australia (Hallac et al. 2012; Demers et al. 2013; Unsworth et al. 2017). Also, some impacts may be more relevant for one habitat type than another within the same region, such as commercial plant harvesting in Atlantic Canada rockweed beds (DFO 2013). Furthermore, our human impact metric does not consider climate change related stressors (i.e., increased sea surface temperature and sea level rise), which impact seagrass through loss of suitable habitat and direct effects on survival (Waycott et al. 2007; Olsen et al. 2012; Valle et al. 2014). Large-scale climate change impacts were not relevant for our comparison of impact scores in seagrass beds across Atlantic Canada but could be included for spatially broader applications to compare different countries or ocean basins.
Application to management and conservation
Our metric is valuable for conservation planning by highlighting areas of low impact and high naturalness, which is a conservation priority for marine protected area placement in Canada and around the world (DFO 2004; Yamakita et al. 2015). For example, in Atlantic NS our impact metric highlights three seagrass beds in the Eastern Shore Archipelago with a high degree of coastal land protection and among the lowest impact scores relative to other beds in this region (Fig. 5; Tables S1 and S2). This provides evidence for the Eastern Shore Archipelago as an ideal candidate for marine protection given the low human impact and high naturalness at both the bay scale and in close proximity to a valuable biogenic habitat (DFO 2019). By assessing impacts in both the terrestrial and marine realms our impact metric will also be valuable for land–sea conservation planning by evaluating coastal ecosystems adjacent to protected land (Álvarez-Romero et al. 2015). For example, in Atlantic NS, Port Joli has high coastal land protection and low riparian land alteration as well as overall low human impact scores relative to other bays in the region (Fig. 5; Tables S1 and S2). In the Gulf of St. Lawrence, Kouchibouguac and St. Louis de Kent in New Brunswick (Fig. 6; Table S3) both have 100% coastal land protection because of the presence of a National Park and also low overall impact scores. Thus, these coastal ecosystems adjacent to protected land would be highly valuable to include in land–sea conservation plans.
The comprehensive, standardized, and multi-scale nature of our metric can also assist in developing management strategies for individual or multiple human impacts at specific sites or for entire coastal bays. For example, in Atlantic NS, Second Peninsula ranks among the most highly impacted sites (Fig. 5; Tables S1 and S2) with nitrogen loading close to the threshold detrimental to seagrass cover (Latimer and Rego 2010), high levels of land use that exceed the threshold for risk of ecosystem degradation in receiving waters (WWF 2017), poor water quality, and no coastal land conservation. Thus, our impact metric identifies risks from multiple human impacts that could be addressed by specific management strategies to reduce or mitigate impacts.
Some human impacts can also be linked to key mechanisms of seagrass growth and survival. For example, light availability is a key determinant of seagrass condition (Hauxwell et al. 2003); thus, knowledge of which seagrass beds are subject to multiple light limiting impacts will be beneficial for management. In the Gulf, our metric identified several seagrass beds at risk from multiple forms of light limitation. For example, nitrogen loading in Bedeque Bay in PEI is among the highest in the Gulf region (Table S3), and one seagrass bed in Bedeque Bay also has among the highest impact scores for overwater structures and riparian land alteration (Table S4). The combination of bay- and local-scale impacts specifically influencing light availability suggests that this bed may be an ideal candidate for management actions targeted at maintaining water clarity, such as reductions in land-derived nutrient inputs (Leschen et al. 2010; Greening et al. 2014; Lefcheck et al. 2018), removal of overwater structures (Thom et al. 2005; Rehr et al. 2014), and conservation of coastal land (Stoms et al. 2005; Álvarez-Romero et al. 2015).
Caveats and the way forward
One current limitation of our metric is that temporal variation could only be accounted for with some impacts (nutrient pollution, invasion extent, and water quality), which represent 10-year averages. The remaining impacts reflect intensity at the present time. This may be problematic if impacts have been historically higher but are currently reduced through management strategies. Inclusion of temporal impact data would be useful to evaluate the response time of biogenic habitats to impacts, and their increases or reductions, and could be included in future applications where relevant data are available.
Our application of the human impact metric is also limited by the disparity in the spatial distribution of seagrass sites among regions (n = 163 and n = 17 for Gulf and Scotian Shelf regions, respectively). Seagrass beds included in our study were those sampled in summer field surveys over the past decade. Inclusion of sites from a large-scale monitoring program in the Gulf region (e.g., Community Aquatic Monitoring Program; Weldon et al. 2009) resulted in a much larger sample size relative to Atlantic NS, where data were compiled from small, individual research projects. The larger dataset in the Gulf region highlighted the importance of within-bay variability, which could not be assessed for the Scotian Shelf region. Further application of our human impact metric as more studies are conducted would be beneficial to fully map the distribution of anthropogenic stressors in Atlantic Canada.
The human impacts included in our metric were chosen based on a comprehensive literature review of human threats to coastal bays and seagrass beds in Atlantic Canada and for which comparable data were available across the region. Lack of the latter prevented the inclusion of a few impacts, which could be added in future applications of our metric once comparable data become available. For example, the invasion extent of European green crabs were not included in our metric as we were unable to obtain quantitative estimates of green crab densities for each of our sites. Also, the data used in our measure of invasion extent represent non-native tunicate presence/absence in or nearby each bay as opposed to growth on seagrass shoots. We assume that increased presence of non-native tunicates in bays that seagrass inhabit will increase the likelihood of fouling on seagrass shoots; however, not enough quantitative data currently exist to verify this for our study sites. In the future, our measure of invasion extent could be improved once green crab monitoring data throughout Atlantic Canada becomes publicly available and if further monitoring of tunicate biofouling on seagrass is conducted. Similarly, other human impacts, such as specific chemical pollution from industrial or municipal point sources could be added to our metric, as well as more refined estimates of boating and fishing activities that directly or indirectly affect seagrass beds or other biogenic habitats.
Finally, further work is necessary to allow the individual and combined impacts to be ranked according to importance for seagrass health. An understanding of the relative importance of each impact will allow the calculation of cumulative scores based on vulnerability weightings, producing one overall metric that is easily interpretable and useful for management and conservation planning.
Conclusions
Given that coastal ecosystems are highly susceptible to human activities, the framework we have developed for quantifying human impacts is valuable for conservation and management planning. Our impact metric allows the finer-scale dynamics of anthropogenic stressors in coastal ecosystems to be explored by uniquely focusing on stressors relevant for coastal biogenic habitats across multiple spatial scales. Our metric can be applied to coastal ecosystems worldwide and can be used to prioritize areas for protection and management by identifying areas of low and high impact, highlighting prominent impacts within and across regions, and describing within-bay variation. The application of our human impact metric to seagrass beds across Atlantic Canada reveals a gradient of human impacts and identifies several bays and seagrass beds that may be good candidates for protection based on high naturalness, and others at risk for future degradation if management strategies are not implemented. Our results can also be used to apply the impact metric to other biogenic habitats in Atlantic Canada and as a baseline of human activities for future comparisons.
Acknowledgments
This research was sponsored by the NSERC Canadian Healthy Oceans Network and its Partners: Department of Fisheries and Oceans Canada and INREST (representing the Port of Sept-Îles and City of Sept-Îles). We would like to thank Marc Skinner, Renee Bernier, Andrea Locke, Monica Boudreau, and the DFO Community Aquatic Monitoring Program for seagrass location data. Thanks to the Canadian Shellfish Sanitation Program for supplying water quality data, the DFO Aquatic Invasive Species Program for supplying invasive species data, Adam Cook for supplying lobster fishing data, and Elizabeth Nagel for assistance with data collection.
References
Álvarez-Romero JG, Pressey RL, Ban NC, and Brodie J. 2015. Advancing land-sea conservation planning: integrating modelling of catchments, land-use change, and river plumes to prioritise catchment management and protection. PLoS ONE, 10: e0145574.
Anderson SC, Mills Flemming J, Watson R, and Lotze HK. 2011. Rapid global expansion of invertebrate fisheries: trends, drivers, and ecosystem effects. PLoS ONE, 6: e14735.
Arasamuthu A, Mathews G, and Patterson Edward JK. 2017. Spatial differences in bacterial and water quality parameters in seagrass meadows of Tuticorin Coast, Gulf of Mannar, southeastern India. Journal of Aquatic Biology and Fisheries, 5: 1–10.
Ban N, and Alder J. 2008. How wild is the ocean? Assessing the intensity of anthropogenic marine activities in British Columbia, Canada. Aquatic Conservation: Marine and Freshwater Ecosystems, 18: 55–85.
Bastien-Daigle S, Hardy M, and Robichaud G. 2007. Habitat management qualitative risk assessment: water column oyster aquaculture in New Brunswick. Canadian Technical Report of Fisheries and Aquatic Sciences 2728. Department of Fisheries and Oceans, Ottawa, Ontario. vii + 72 p.
Beazley L, Kenchington E, and Lirette C. 2017. Species distribution modelling and kernel density analysis of benthic ecologically and biologically significant areas (EBSAs) and other benthic fauna in the Maritimes Region. Canadian Technical Report of Fisheries and Aquatic Sciences 3204. Department of Fisheries and Oceans, Ottawa, Ontario.
Beck MW, Brumbaugh RD, Airoldi L, Carranza A, Coen LD, Crawford C, et al. 2011. Oyster reefs at risk and recommendations for conservation, restoration, and management. BioScience, 61: 107–116.
Bilkovic DM, Roggero M, Hershner CH, and Havens KH. 2006. Influence of land use on microbenthic communities in nearshore estuarine habitats. Estuaries and Coasts, 29: 1185–1195.
Blake RE, Duffy JE, and Richardson JP. 2014. Patterns of seagrass community response to local shoreline development. Estuaries and Coasts, 37:1549–1561.
Braga RR, Gomez-Aparicio L, Heger T, Vitule JRS, and Jeschke JM. 2018. Structuring evidence for invasional meltdown: broad support but with biases and gaps. Biological Invasions, 20: 923–936.
Breininger DR, Breininger RD, and Hall CR. 2016. Effects of surrounding land use and water depth on seagrass dynamics relative to a catastrophic algal bloom. Conservation Biology, 31:67–75.
Brush ML, and Nixon SW. 2002. Direct measurements of light attenuation by epiphytes on eelgrass Zostera marina. Marine Ecology Progress Series, 238: 73–79.
Bryce SA, Lomnicky GA, and Kaufmann PR. 2010. Protecting sediment-sensitive aquatic species in mountain streams through the application of biologically based streambed sediment criteria. Journal of the North American Benthological Society, 29: 657–672.
Bugden G, Jiang Y, van den Heuvel M, Vandermeulen H, MacQuarrie K, Crane C, et al. 2014. Nitrogen loading criteria for estuaries in Prince Edward Island. Canadian Technical Report of Fisheries and Aquatic Sciences 3066. Department of Fisheries and Oceans, Ottawa, Ontario.
Burgin S, and Hardiman N. 2011. The direct physical, chemical and biotic impacts on Australian coastal waters due to recreational boating. Biodiversity and Conservation, 20: 683–701.
Coll M, Schmidt A, Romanuk T, and Lotze HK. 2011. Food-web structure of seagrass communities across different spatial scales and human impacts. PLoS ONE, 6: e22591.
Comeau LA. 2013. Suspended versus bottom oyster culture in eastern Canada: comparing stocking densities and clearance rates. Aquaculture, 410: 57–65.
CSSP. 2016. Canadian Shellfish Sanitation Program [online]: Available from inspection.gc.ca/food/fish-and-seafood/shellfish-sanitation/eng/1299826806807/1299826912745.
Cullain N, McIver R, Schmidt AL, Milewski I, and Lotze HK. 2018a. Impacts of organic enrichment from finfish aquaculture on seagrass beds and associated macroinfaunal communities in Atlantic Canada. PeerJ, 6: e5630.
Cullain N, McIver R, Schmidt AL, and Lotze HK. 2018b. Spatial variation of macroinfaunal communities associated with Zostera marina beds across three biogeographic regions in Atlantic Canada. Estuaries and Coasts, 41: 1381–1396.
Cullen-Unsworth LC, and Unsworth RKF. 2016. Strategies to enhance the resilience of the world’s seagrass meadows. Journal of Applied Ecology, 53: 967–972.
Demers M-CA, Davis AR, and Knott NA. 2013. A comparison of the impact of ‘seagrass-friendly’ boat mooring systems on Posidonia australis. Marine Environmental Research, 83:54–62.
Department of Fisheries and Oceans (DFO) Canada. 2004. Evaluation of site selection methodologies for use in marine protected area network design. Canadian Science Advisory Secretariat Science Advisory Report 2004/082. Department of Fisheries and Oceans, Ottawa, Ontario.
Department of Fisheries and Oceans (DFO) Canada. 2007. Ecologically and biologically significant areas (EBSA) in the Estuary and Gulf of St. Lawrence: identification and characterization. Canadian Science Advisory Secretariat Science Advisory Report 2007/016. Department of Fisheries and Oceans, Ottawa, Ontario.
Department of Fisheries and Oceans (DFO) Canada. 2009a. Does eelgrass (Zostera marina) meet the criteria as an ecologically significant species? Canadian Science Advisory Secretariat Science Advisory Report 2009/018. Department of Fisheries and Oceans, Ottawa, Ontario.
Department of Fisheries and Oceans (DFO) Canada. 2009b. Development of a framework and principles for the biogeographic classification of Canadian marine areas. Canadian Science Advisory Secretariat Science Advisory Report 2009/056. Department of Fisheries and Oceans, Ottawa, Ontario.
Department of Fisheries and Oceans (DFO) Canada. 2013. Assessment of information on Irish moss, rockweed, and kelp harvests in Nova Scotia. Canadian Science Advisory Secretariat Science Advisory Report 2013/004. Department of Fisheries and Oceans, Ottawa, Ontario.
Department of Fisheries and Oceans (DFO) Canada. 2018. Design strategies for a network of marine protected areas in the Scotian shelf bioregion. Canadian Science Advisory Secretariat Science Advisory Report 2018/006. Department of Fisheries and Oceans, Ottawa, Ontario.
Department of Fisheries and Oceans (DFO) Canada. 2019. Biophysical and ecological overview of the Eastern Shore Islands Area of Interest (AOI). Canadian Science Advisory Secretariat Science Advisory Report 2019/025. Department of Fisheries and Oceans, Ottawa, Ontario.
Duarte CM. 2002. The future of seagrass meadows. Environmental Conservation, 29: 192–206.
Fresh KL, Wyllie-Echeverria T, Wyllie-Echeverria S, and Williams BW. 2006. Using light-permeable grating to mitigate impacts of residential floats on eelgrass Zostera marina L. in Puget Sound, Washington. Ecological Engineering, 28: 354–362.
Garbary DJ, Miller AG, Williams J, and Seymour NR. 2014. Drastic decline of an extensive eelgrass bed in Nova Scotia due to the activity of the invasive green crab (Carcinus maenas). Marine Biology, 161: 3–15.
Google Inc. 2018. Google Earth Pro (version 7.3.2.5491) [Software]. Available from: google.com/earth/versions/.
Grech A, Chartrand-Miller K, Erftemeijer P, Fonseca M, McKenzie L, Rasheed M, et al. 2012. A comparison of threats, vulnerabilities and management approaches in global seagrass bioregions. Environmental Research Letters, 7: 1–8.
Greening H, Janicki A, Sherwood ET, Pribble R, and Johansson JOR. 2014. Ecosystem responses to long-term nutrient management in an urban estuary: Tampa Bay, Florida, USA. Estuarine, Coastal, and Shelf Science, 151: A1–A16.
Guyondet T, Sonier R, and Comeau LA. 2013. A spatially explicit seston depletion index to optimize shellfish culture. Aquaculture Environment Interactions, 4: 175–186.
Hallac D, Sadle J, Pearlstine LG, Herling F, and Shinde D. 2012. Boating impacts to seagrass in Florida Bay, Everglades National Park, Florida, USA: links with physical and visitor-use factors and implications for management. Marine and Freshwater Research, 63: 1117–1128.
Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D’Agrosa C, et al. 2008. A global map of human impact on marine ecosystems. Science, 319: 948–952.
Hauxwell J, Cebrián J, and Valiela I. 2003. Eelgrass Zostera marina loss in temperate estuaries: Relationship to land-derived nitrogen loads and effect of light limitation imposed by algae. Marine Ecology Progress Series, 247: 59–73.
Hemminga MA, and Duarte CM. 2000. Seagrass ecology. Cambridge University Press, Cambridge, UK.
Hitchcock JK, Courtenay SC, Coffin MRS, Pater CC, and van den Heuvel MR. 2017. Eelgrass bed structure, leaf nutrient, and leaf isotope responses to natural and anthropogenic gradients in estuaries of the southern Gulf of St. Lawrence, Canada. Estuaries and Coasts, 40: 1653–1665.
Huang J, Huang Y, Pontius RG Jr, and Zhang Z. 2015. Geographically weighted regression to measure spatial variations in correlations between water pollution versus land use in a coastal watershed. Ocean and Coastal Management, 103: 14–24.
Hughes AR, Williams SL, Duarte CM, Heck KL Jr, and Waycott M. 2009. Associations of concern: declining seagrasses and threatened dependent species. Frontiers in Ecology and the Environment, 7: 242–246.
Iacarella JC, Adamczyk E, Bowen D, Chalifour L, Eger A, Heath W, et al. 2018. Anthropogenic disturbance homogenizes seagrass fish communities. Global Change Biology, 24: 1904–1918.
Krumhansl KA, Okamoto DK, Rassweiler A, Novak M, Bolton JJ, Cavanaugh KC, et al. 2016. Global patterns of kelp forest change over the past half-century. Proceedings of the National Academy of Sciences of the USA, 113: 13785–13790.
Lamb JB, van de Water JAJM, Bourne DG, Altier C, Hein MY, Fiorenza EA, et al. 2017. Seagrass ecosystems reduce exposure to bacterial pathogens of humans, fishes, and invertebrates. Science, 355: 731–733.
Latimer JS, and Rego SA. 2010. Empirical relationship between eelgrass extent and predicted watershed-derived nitrogen loading for shallow New England estuaries. Estuarine, Coastal and Shelf Science, 90: 231–240.
Lefcheck JS, Orth RJ, Dennison WC, Wilcox DJ, Murphy RR, Keisman J, et al. 2018. Long-term nutrient reductions lead to the unprecedented recovery of a temperate coastal region. Proceedings of the National Academy of Sciences of the USA, 115: 3658–3662.
Leitão RP, Zuanon J, Mouillot D, Leal CG, Hughes RM, Kaufmann PR, et al. 2018. Disentangling the pathways of land use impacts on the functional structure of fish assemblages in Amazon streams. Ecography, 41: 219–232.
Lerberg SB, Holland AF, and Sanger DM. 2000. Responses of tidal creek macrobenthic communities to the effects of watershed development. Estuaries, 23: 838–853.
Leschen AS, Ford KH, and Evans NT. 2010. Successful eelgrass (Zostera marina) restoration in a formerly eutrophic estuary (Boston Harbor) supports the use of a multifaceted watershed approach to mitigating eelgrass loss. Estuaries and Coasts, 33: 1340–1354.
Lotze HK, Lenihan HS, Bourque BJ, Bradbury RH, Cooke RG, Kay MC, et al. 2006. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science, 312: 1806–1809.
Lowen JB, and DiBacco C. 2017. Distributional changes in a guild of non-indigenous tunicates in the NW Atlantic under high-resolution climate projections. Marine Ecology Progress Series, 570: 173–186.
McIver R, Milewski I, and Lotze HK. 2015. Land use and nitrogen loading in seven estuaries along the southern Gulf of St. Lawrence, Canada. Estuarine, Coastal and Shelf Science, 165: 137–148.
McIver R, Milewski I, Loucks R, and Smith R. 2018. Estimating nitrogen loading and far-field dispersal potential from background sources and coastal finfish aquaculture: a simple framework and case study in Atlantic Canada. Estuarine, Coastal and Shelf Science, 205: 46–57.
McIver R, Schmidt AL, Cullain N, and Lotze HK. 2019. Linking estimates of nitrogen loading and watershed characteristics to eelgrass bed structure and eutrophication symptoms across 7 bays in Atlantic Canada. Marine Environmental Research, 144: 141–153.
Milewski I, Loucks RH, Fisher B, Smith RE, McCain JSP, and Lotze HK. 2018. Sea-cage aquaculture impacts market and berried lobster (Homarus americanus) catches. Marine Ecology Progress Series, 598: 85–97.
Moore A, Vercaemer B, DiBacco C, Sephton D, and Ma K. 2014. Invading Nova Scotia: first records of Didemnum vexillum Kott, 2002 and four more non-indigenous invertebrates in 2012 and 2013. BioInvasions Records, 3: 225–234.
Murray CC, Agbayani S, Alidini HM, and Ban NC. 2015. Advancing marine cumulative effects mapping: an update in Canada’s Pacific waters. Marine Policy, 58: 71–77.
Nagel EJ, Murphy G, Wong MC, and Lotze HK. 2018. Nitrogen loading rates for twenty-one seagrass inhabited bays in Nova Scotia, Canada. Canadian Technical Report of Fisheries and Aquatic Sciences 3260. v + 37 p.
Olsen YS, Sánchez-Camacho M, Marbà N, and Duarte CM. 2012. Mediterranean seagrass growth and demography responses to experimental warming. Estuaries and Coasts, 35: 1205–1213.
Orth RJ, Carruthers TJB, Dennison WC, Duarte CM, Fourqurean JW, Heck KL Jr, et al. 2006. A global crisis for seagrass ecosystems. BioScience, 56: 987–996.
Palmer T. 2018. Nitrogen loading rates for Prince Edward Island estuaries and effects on eelgrass (Zostera marina) structural characteristics. Honours thesis, Dalhousie University, Halifax, Nova Scotia. p. 13–16.
Parker J, Epifanio J, Casper A, and Cao Y. 2016. The effects of improved water quality on fish assemblages in a heavily modified large river system. River Research and Applications, 32: 992–1007.
Powell KI, Chase JM, and Knight TM. 2013. Invasive plants have scale-dependent effects on diversity by altering species-area relationships. Science, 339: 316–318.
Quiros TEA, 2016. Linking terrestrial and marine protected areas at the coastal interface. Ph.D. thesis, University of California, Santa Cruz, Santa Cruz, California, USA. p. 40–43.
Quiros TEA, Croll D, Tershy C, Fortes MD, and Raimondi P. 2017. Land use is a better predictor of tropical seagrass condition than marine protection. Biological Conservation, 209: 454–463.
R Core Team. 2015. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria [online]: Available from R-project.org/.
Rehr AP, Williams GD, Tolimeieri N, and Levin PS. 2014. Impacts of terrestrial and shoreline stressors on eelgrass in Puget Sound: an expert elicitation. Coastal Management, 42: 246–262.
Schmidt AL, Wysmyk JKC, Craig SE, and Lotze HK. 2012. Regional-scale effects of eutrophication on ecosystem structure and services of seagrass beds. Limnology and Oceanography, 57: 1389–1402.
Sephton D, Vercaemer B, Silva A, Stiles L, Harris M, and Godin K. 2017. Biofouling monitoring for aquatic invasive species (AIS) in DFO Maritimes Region (Atlantic shore of Nova Scotia and southwest New Brunswick): May–November, 2012–2015. Canadian Technical Report of Fisheries and Aquatic Sciences 3158. Department of Fisheries and Oceans, Ottawa, Ontario. ix + 172 p.
Shelton AO, Francis TB, Feist BE, Williams GD, Lindquist A, and Levin PS. 2017. Forty years of seagrass population stability and resilience in an urbanizing estuary. Journal of Ecology, 105: 458–470.
Short FT, and Wyllie-Echeverria S. 1996. Natural and human-induced disturbance of seagrasses. Environmental Conservation, 23: 17–27.
Simberloff D, and Von Holle B. 1999. Positive interactions of nonindigenous species: invasional meltdown? Biological Invasions, 1: 21–32.
Simpson SD, Radford AN, Nedelec SL, Ferrari MCO, Chivers DP, Mccormick MI, et al. 2016. Anthropogenic noise increases fish mortality by predation. Nature Communications, 7: 10544.
Skinner MA, Courtenay SC, and McKindsey CW. 2013. Reductions in distribution, photosynthesis, and productivity of eelgrass Zostera marina associated with oyster Crassostrea virginica aquaculture. Marine Ecology Progress Series, 486: 105–119.
Spalding MD, Fox HE, Allen GR, Davidson N, Ferdaña ZA, Finlayson M, et al. 2007. Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. BioScience, 57: 573–583.
Statistics Canada. 2017. 2011 Census of Canada: Topic-based tabulations, Structural type of dwelling and collectives [online]: Available from statcan.gc.ca/census-recensement/2011/dp-pd/tbt-tt/Index-eng.cfm.
Stoms DM, Davis FW, Andelman SJ, Carr MH, Gaines SD, Halpern BS, et al. 2005. Integrated coastal reserve planning: Making the land-sea connection. Frontiers in Ecology and the Environment, 3: 429–436.
Thom RM, Williams GW, and Diefenderfer HL. 2005. Balancing the need to develop coastal areas with the desire for an ecologically functioning coastal environment: is net ecosystem improvement possible? Restoration Ecology, 13: 193–203.
Thom RM, Buenau KE, Judd C, and Cullinan VI. 2011. Eelgrass (Zostera marina) stressors in Puget Sound. US Department of Energy, Washington, D.C., USA.
Thrush SF, Lawrie SM, Hewitt JE, and Cummings VJ. 1999. The problem of scale: uncertainties and implications for soft-bottom marine communities and the assessment of human impacts. Biogeochemical Cycling and Sediment Ecology, 59: 195–210.
Uhrin AV, Fonesca MS, and DiDomenico GP. 2005. Effect of Caribbean spiny lobster traps on seagrass beds of the Florida Keys National Marine Sanctuary: damage assessment and evaluation of recovery. American Fisheries Society Symposium, 41: 579–588.
Unsworth RKF, Williams B, Jones BL, and Cullen-Unsworth LC. 2017. Rocking the boat: damage to eelgrass by swinging boat moorings. Frontiers in Plant Science, 8: 1309.
Uriarte M, Yackulic CB, Lim Y, and Arce-Nazario JA. 2011. Influence of land use on water quality in a tropical landscape: a multi-scale analysis. Landscape Ecology, 26: 1151–1164.
Valiela I, Collins G, Kremer J, Lajtha K, Geist M, Seely B, et al. 1997. Nitrogen loading from coastal watersheds to receiving estuaries: New method and application. Ecological Applications, 7: 358–380.
Valle M, Chust G, del Campo A, Wisz MS, Olsen SM, Garmendia JM, et al. 2014. Projecting future distribution of the seagrass Zostera noltii under global warming and sea level rise. Biological Conservation, 170: 74–85.
Van Katwijk MM, van der Welle MEW, Lucassen ECHET, Vonk JA, Christianen MJA, Kiswara W, et al. 2011. Early warning indicators for river nutrient and sediment loads in tropical seagrass beds: a benchmark from a near-pristine archipelago in Indonesia. Marine Pollution Bulletin, 62: 1512–1520.
Vance A. 2014. Applying an ecosystem-based risk management approach to the relationship between eelgrass beds and oyster aquaculture at multiple spatial scales in eastern New Brunswick, Atlantic Canada. Master of Marine Management thesis, Dalhousie University, Halifax, Nova Scotia. p. 34–37.
Vercaemer B, Sephton D, Clément P, Harman A, Stewart-Clark S, and Dibacco C. 2015. Distribution of the non-indigenous colonial ascidian Didemnum vexillum (Kott, 2002) in the Bay of Fundy and on offshore banks, eastern Canada. Managing Biological Invasions, 6: 385–394.
Waycott M, Collier C, Mcmahon K, Ralph P, Mckenzie L, Udy J, et al. 2007. Vulnerability of seagrasses in the Great Barrier Reef to climate change. In Climate Change and the Great Barrier Reef: A Vulnerability Assessment. Edited by J.E. Johnson and P.A. Marshal. Great Barrier Reef Marine Park Authority and Australian Greenhouse Office, Townsville, Australia. pp. 193–299.
Waycott M, Duarte CM, Carruthers TJB, Orth RJ, Dennison WC, Olyarnik S, et al. 2009. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proceedings of the National Academy of Science of the USA, 106: 12377–12381.
Weldon J, Courtenay S, and Garbary D. 2009. The community aquatic monitoring program (CAMP) for measuring marine environmental health in coastal waters of the southern Gulf of St. Lawrence: 2007 overview. Canadian Technical Report of Fisheries and Aquatic Sciences 2825. viii + 75 p.
Wickham H. 2016. ggplot2: elegant graphics for data analysis. Springer-Verlag, New York, USA.
Williams SL. 2007. Introduced species in seagrass ecosystems: status and concerns. Journal of Experimental Marine Biology and Ecology, 350: 89–110.
Wong MC. 2018. Secondary production of macrobenthic communities in seagrass (Zostera marina, eelgrass) beds and bare soft sediments across differing environmental conditions in Atlantic Canada. Estuaries and Coasts, 41: 536–548.
Wong MC, and Vercaemer B. 2012. Effects of invasive colonial tunicates and a native sponge on the growth, survival, and light attenuation of eelgrass (Zostera marina). Aquatic Invasions, 7: 315–326.
World Wildlife Fund (WWF). 2017. A national assessment of Canada’s freshwater watershed reports. WWF Canada, Toronto, Canada.
Worm B, and Lotze HK. 2006. Effects of eutrophication, grazing, and algal blooms on rocky shores. Limnology and Oceanography, 51: 569–579.
Yamakita T, Yamamoto H, Nakaoka M, Yamano H, Fujukura K, Hidaka K, et al. 2015. Identification of important marine areas around the Japanese Archipelago: establishment of a protocol for evaluating a broad area using ecologically and biologically significant areas selection criteria. Marine Policy, 51: 136–147.
Yates AG, and Bailey RC. 2010. Improving the description of human activities potentially affecting rural stream ecosystems. Landscape Ecology, 25: 371–382.
Supplementary materials
Supplementary Material 1 (DOCX / 1.87 MB)
- Download
- 1.87 MB
Supplementary Material 2 (CSV / 12.9 KB)
- Download
- 12.93 KB
Supplementary Material 3 (CSV / 21.6 KB)
- Download
- 21.65 KB
Supplementary Material 4 (DOCX / 16 KB)
- Download
- 16.03 KB
Information & Authors
Information
Published In

FACETS
Volume 4 • Number 1 • June 2019
Pages: 210 - 237
Editor: Sophia Johannessen
History
Received: 30 October 2018
Accepted: 9 January 2019
Version of record online: 14 June 2019
Copyright
© 2019 Murphy et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper and in the Supplementary Material.
Key Words
Sections
Subjects
Plain Language Summary
Researchers create report card for quantifying human impacts on coastal ecosystems
Authors
Author Contributions
All conceived and designed the study.
GEPM performed the experiments/collected the data.
All analyzed and interpreted the data.
All contributed resources.
All drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Grace E.P. Murphy, Melisa C. Wong, and Heike K. Lotze. 2019. A human impact metric for coastal ecosystems with application to seagrass beds in Atlantic Canada. FACETS.
4(1): 210-237. https://doi.org/10.1139/facets-2018-0044
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Loss, resilience and recovery of kelp forests in a region of rapid ocean warming
2. Hunting and seagrass affect fall stopover Canada goose distribution in eastern Canada
3. Coastal community perceptions of eelgrass in Atlantic Canada: Considerations for management
4. Atlantic Coast Rivers of Canada
5. Cumulative effects of long-term anthropogenic impacts on resident fish communities in subarctic fishing harbours
6. Fine-scale ensemble species distribution modeling of eelgrass (Zostera marina) to inform nearshore conservation planning and habitat management
7. An Evaluation Framework for Risk of Coastal Marine Ecological Diversity Loss From Land-Based Impacts
8. Incorporating anthropogenic thresholds to improve understanding of cumulative effects on seagrass beds
9. Variability of bio-optical properties in nearshore waters of the estuary and Gulf of St. Lawrence: Absorption and backscattering coefficients
10. Anthropogenic nitrogen loading and risk of eutrophication in the coastal zone of Atlantic Canada
11. The Evidence of Seagrass Environmental Support for Local People’s Economic on the South Coast of Lombok Island
12. Reproductive ecology of Zostera marina L. (eelgrass) across varying environmental conditions
13. Interactions between finfish aquaculture and American lobster in Atlantic Canada
14. Inferring the potential for nitrogen toxicity on seagrass in the vicinity of an aquaculture site using mathematical models
15. Multiple Metrics of Temperature, Light, and Water Motion Drive Gradients in Eelgrass Productivity and Resilience
16. Using Landsat Time-Series to Monitor and Inform Seagrass Dynamics: A Case Study in the Tabusintac Estuary, New Brunswick, Canada
17. From coast to coast to coast: ecology and management of seagrass ecosystems across Canada
18. Aquaculture and eelgrass Zostera marina interactions in temperate ecosystems
19. Branching Algorithm to Identify Bottom Habitat in the Optically Complex Coastal Waters of Atlantic Canada Using Sentinel-2 Satellite Imagery
20. Response and Recovery of Eelgrass (Zostera marina) to Chronic and Episodic Light Disturbance
21. Eelgrass (Zostera marina) and benthic habitat mapping in Atlantic Canada using high-resolution SPOT 6/7 satellite imagery
22. Ecosystem-based management of seaweed harvesting
23. Climate change projections reveal range shifts of eelgrass Zostera marina in the Northwest Atlantic