Temperature-dependent infection of freshwater turtle hatchlings, Emydura macquarii krefftii, inoculated with a ranavirus isolate (Bohle iridovirus, Iridoviridae)
Abstract
Fish, amphibians, and reptiles exhibit temperature-dependent ranaviral disease. We performed an experimental infection at four different environmental temperatures (16, 22, 28, and 34 °C) to investigate the effect of temperature on ranaviral infection in Krefft’s turtle (Emydura macquarii krefftii). Infection rates and viral loads were determined by quantitative polymerase chain reaction to detect ranaviral DNA in liver samples at 21 d postexposure. The rate of infection differed across the temperature treatment groups. Infection rates were 44%, 90%, 60%, and 10% for the 16, 22, 28, and 34 °C temperature groups, respectively. Highest viral load was observed in the 28 °C temperature group, and there was a statistically significant difference in viral load between the 16 and 28 °C temperature groups (p = 0.027). Based on the results of this study, the temperature of maximal infection rate for ranaviral infection in Krefft’s river turtles is estimated to be 23.2 °C (SD = 4.5). The findings of this study can inform management decisions in terms of disease control and treatment and form a platform for modelling disease outbreaks.
Introduction
Ranaviruses are pathogens of ectothermic vertebrates like reptiles. Because reptile physiology is strongly influenced by the temperature of their surrounding environment, so is the pathogenesis of ranaviral infections. The environmental temperature-dependent physiology includes many aspects of the ectothermic immune system (both adaptive and innate) and, while immune function is often severely reduced at lower temperatures, many immune components have optimal temperatures beyond which the efficacy diminishes (Zimmerman et al. 2010).
As with the ectothermic immune response, the replication rate of ranaviruses is also linked to temperature (Ariel et al. 2009). It has been concluded that the decrease in mortality at higher temperatures (>25 °C) is associated with ranaviruses’ inability to replicate effectively above 32 °C in cell culture (Chinchar et al. 2009; Allender et al. 2013). However, it is difficult to differentiate the degree to which temperature-dependent pathogenesis is a result of the effect of temperature on the replication of the virus or on the immune system or other physiological components of the ectothermic host (Brunner et al. 2015).
Given the effects of temperature on the ectothermic immune system and viral replication, ranaviral disease is, in part, determined by the environmental temperature at which infection occurs. Wild outbreaks of ranaviruses are often seasonal and theoretically driven by temperature (Hall et al. 2018). Challenge studies have shown that changes in environmental temperature can affect pathogenesis in exposed animals. Ranavirus-infected turtles exhibit temperature-dependent disease (Allender et al. 2013, 2018). Similarly, fish and amphibians exhibit temperature-dependent ranaviral disease (Whittington and Reddacliff 1995; Rojas et al. 2005; Ariel and Jensen 2009; Jensen et al. 2011; Brunner et al. 2015; Brand et al. 2016).
As temperature can influence ranaviral disease, temperature therapy (exposing animals to environmental temperatures that are associated with reduced disease) may be a cheap and effective method for treating acute ranaviral disease in captive animals if the optimal temperature for survival is known. High environmental temperatures may effectively force the animals into behavioural fever, increasing their immune response and reducing viral replication, which subsequently results in reduced mortality. Behavioural fever is the process by which a diseased ectotherm (e.g., reptile) actively brings about a febrile state through the behavioural selection of higher environmental temperatures (Monagas and Gatten 1983; do Amaral et al. 2002; Merchant et al. 2007). Infected or antigen-challenged ectotherms will generally select an environmental temperature a few degrees higher than control groups, and it is thought that this temperature is correlated with the optimal immune response of the animal (Spellerberg 1972; Monagas and Gatten 1983; Merchant et al. 2007). There is evidence that toads perform behavioural fever to fight ranaviral infections (Sauer et al. 2019). Thus, knowing the optimal temperature of infection, it will be possible to make recommendations of the optimal temperature for temperature therapy and the best temperatures for basking sites in enclosures to ensure behavioural fever is possible.
Understanding the effect of temperature on infection also has important ramifications for understating wild disease dynamics. Knowing the rage of temperatures that infection can occur at and the optimal temperature for infection, it may be possible to identify at-risk populations or predict temperature-driven epizootics. As the climate continues to change it is important to understand how variation in temperature may affect aspects of ranaviral disease.
Australia is home to ranaviruses, frog virus 3 (FV3) and epizootic hematopoietic necrosis virus, and susceptible ectothermic vertebrates such as the Krefft’s river turtle (Emydura macquarii krefftii) (Ariel et al. 2015; Chinchar et al. 2018). Krefft’s river turtles belong to the Pleurodira suborder of turtles, which are distinct from the native turtles of Asia, Europe, and North America and may be a useful model for other Pleurodirid turtles (of Africa, Australasia, and South America). The effect of ranaviral dose at a single temperature has been reported for Krefft’s hatchlings (Wirth et al. 2019). This study investigates how infection rate with a local ranaviral isolate (Bohle iridovirus; FV3) varies within a range of environmental temperatures representative of topical Australia (16–34 °C).
Methods
All experiments were carried out under a James Cook University (JCU) Animal Ethics permit (A2344) and a Department of Environment and Science Scientific Research Permit (WISP13270413).
Animals
Hatchlings used in this study were obtained following methods described by Wirth et al. (2019). Briefly, wild gravid turtles were induced to lay eggs with an injection of 1 IU 100 g−1 of syntocinon (synthetic oxytocin 10 IU mL−1, Troy laboratories). After approximately 50 d of incubation at 29 °C the eggs hatched, and newly emerged hatchlings were kept at the JCU Turtle Health Research Facility. The hatchlings were housed in tanks with water between 24 and 28 °C and given access to dry areas for basking. The hatchlings were fed commercial turtle pellets (Exo Terra, aquatic turtle food) ad libitum for 10–15 min per day. During and after this husbandry phase, feeding observations and morphometric data were collected from each individual. The average weight of hatchlings used in this study was 9.5 g (Table 1).
Table 1.
| Group | Temperature (°C) | Number of hatchlings | Mean starting weight (g) |
|---|---|---|---|
| Control | 16 | 2* | 10.8 |
| 22 | 3 | 9.4 | |
| 28 | 3 | 9.7 | |
| 34 | 3 | 9.3 | |
| Exposed | 16 | 9* | 9.4 |
| 22 | 10 | 9.5 | |
| 28 | 10 | 9.4 | |
| 34 | 10 | 9.5 |
Note: During the study one animal from both the control and exposed 16 °C temperature groups were removed.
*
The weights of the removed animals are not included in the mean starting weight.
Source of virus
The Bohle iridovirus (BIV) isolate (FV3 strain) used in this study was originally isolated in Townsville, Australia and was produced according to the methods of Maclaine et al. (2018). Briefly, the viral isolate was propagated at 25 °C in fathead minnow cells grown in Dulbecco’s modified eagle medium (DMEM; Thermo Fisher Scientific), supplemented with 100× antibiotic-antimycotic (Thermo Fisher Scientific) and 10% fetal bovine serum (Bovogen Biologicals). Viral titre (TCID50) was determined using the methods described by Reed and Muench (1938). The viral stock was stored at −80 °C until the time of challenge study and then diluted in DMEM to 102.5 TCID50 mL−1.
Experimental design
One week prior to the start of the experiment animals were randomly assigned to control and exposed groups (Table 1) and moved into the infection room. The hatchlings were kept in individual 500 mL plastic container on a slant to create a dry area for basking. The containers were placed, in groups of three or four, in temperature chambers set to 16, 22, 28, or 34 °C with a 12-h light cycle (Greenspan et al. 2016). The hatchlings were given one week to acclimatise to the temperature chambers before the start of the experiment. Husbandry continued as previously described. During the acclimatisation period and throughout the experiment hatchlings were monitored for clinical signs including inappetence, skin lesions, increased basking behaviour, and oedema of the neck and legs as described by Wirth et al. (2019) for ranaviral infection in Krefft’s river turtle hatchlings. The dose rate and time to euthanasia in this experiment was based on findings in Wirth et al. (2019) to allow for sufficient time for the infection to get established and potentially be influenced by temperatures above and below the temperature used in that study, before the samples were collected. On day zero of the experiment all hatchlings in the exposed groups received an intramuscular injection of 102.5 TCID50 mL−1 BIV stock into the hind leg. Injection volume was determined by multiplying the individual’s weight (g) by 2.5 (mean 23.8 μL; range 20.25–30.5 μL). Control animals received an equivalent dose of virus-free DMEM as a placebo.
Endpoint and sample collection
On the day of euthanasia final morphometric data were collected. Hatchlings were randomly selected for euthanisa 21 d post exposure and all hatchlings were euthanized over the following 3 d. Hatchlings were euthanised with a MS-222 overdose according to the methods described by Conroy et al. (2009). A necropsy was performed on each animal using a different set of sterile tools (disinfected with Virkon and washed with detergent). Gloves were changed between animals and surfaces were decontaminated with bleach (4% by volume). A section of liver was collected for quantitative polymerase chain reaction (qPCR) analysis and frozen at −80 °C.
qPCR
DNA was extracted from the thawed liver samples using an ISOLATE II Genomic DNA Kit (Bioline) following the manufacturer’s protocol for DNA extraction from tissues. Viral loads were determined using the ranavirus major capsid protein gene (MCP) gene assay described by Leung et al. (2017). A single copy conserved vertebrate gene, EBF3N, was used to standardise the number of viral copies. A section of the BIV MCP and EBF3N gene were cloned into pGEM-easy vectors following the manufacturer’s protocol (Promega). Plasmids were linearised using a PstI restriction digest (New England Biolabs) and quantified using a QuantiFluor dye system (QuantiFluor, Promega). Absolute standard curves were constructed in triplicate from 108 to 101 copies μL−1 and 107–101 copies μL−1 for the MCP and EBF3N plasmids, respectively. The qPCR was performed on a magnetic induction cycler real-time thermocycler (Applied Biosystems) with the conditions described by Leung et al. (2017). The assays were run as a duplex with the MCP probe on the green channel (FAM probe) and ENF3N on the red channel (Cy5 probe). It was assumed that there is one EBF3N gene per E. m. krefftii genome. Viral loads were determined by dividing the number of MCP copies μL−1 multiplied by two by the number of EBF3N copies μL−1 and are referred to as copies per cell throughout this manuscript.
Statistics
All analysis was performed using the Statsmodels module (0.10.2) and Pandas (0.24.3) and SciPy (1.4.1) libraries in Python 3.6 (McKinney 2010; Seabold and Perktold 2010). A 2 × 4 χ2 contingency table was used to compare proportions of infected animals in the temperature groups. The Kruskal–Wallis H-test for independent samples was used to compare variation in viral loads of infected turtles between temperature groups. Conover’s test of multiple comparisons was used as post hoc test following a significant Kruskal–Wallis H-test. A normal distribution was fit to the portion of infected animals from each temperature group (using mean temperature, Table 2) to estimate the temperature of maximal infection rate and its standard deviation for the temperature range tested.
Table 2.
| Group | Mean | Std | Minimum | Q1 | Median | Q3 | Maximum |
|---|---|---|---|---|---|---|---|
| 16 | 16.24 | 0.72 | 15.9 | 16 | 16.1 | 16.3 | 23.3 |
| 22 | 22.63 | 1.13 | 21.6 | 22 | 22.3 | 22.5 | 27.6 |
| 28 | 27.79 | 1.12 | 22.9 | 27.8 | 28.1 | 28.4 | 29.1 |
| 34 | 34.12 | 0.35 | 32.1 | 33.8 | 34.1 | 34.4 | 35 |
Note: Q1 and Q3 represent the lower and upper quartiles respectively. All data are expressed as degrees Celsius (°C).
Results
Temperatures
The incubators used in this study were effective at maintaining the average environmental temperature within a degree of the target temperature (Table 2).
Deaths
During the study two animals in the 16 °C group were found dead in their enclosures on day 3 and 8 postinoculation. One animal was from the control group and one from the infection group. Following a postmortem, it was determined that both animals had drowned and not died from a ranaviral infection (they were qPCR negative). The animals were removed from the study and not included in any analysis. No other animals died or developed clinical signs severe enough to warrant euthanasia.
Clinical signs
Only two animals developed clinical signs in the timeframe of this experiment. One animal in the 16 °C group developed ocular and nasal lesions 17 d after exposure. One animal in the 22 °C group developed an oral lesion 14 d after exposure. No animals developed skin lesions at the site of inoculation as described by Wirth et al. (2019). Diet consumption appeared to be strongly linked to environmental temperature and there was no clear evidence of in-appetence caused by ranaviral infection (control and infected groups were affected similarly across all temperatures). Both exposed and control hatchlings in the 16 °C group consistently ate little to none of their food and appeared lethargic. The animals in the 34 °C group consistently ate most of the food offered and were active throughout the experiment.
qPCR
Of the 40 turtles infected across all temperatures 50% of them reacted in the qPCR assay for ranaviral DNA in the liver 21 d after exposure. However, the distribution of reactors was not even (Fig. 1). Significantly more turtles (9/10 liver samples reacted in the qPCR assay) were infected in the 22 °C group (χ2 = 13.3; p < 0.01). The lowest infection rate was in the turtles kept at 34 °C, where only 1/10 liver samples reacted in the qPCR assay (10%). The mean and standard deviation of the normal distribution fit to the infection rates was 23.2 and 4.5 °C, respectively.
Fig. 1.
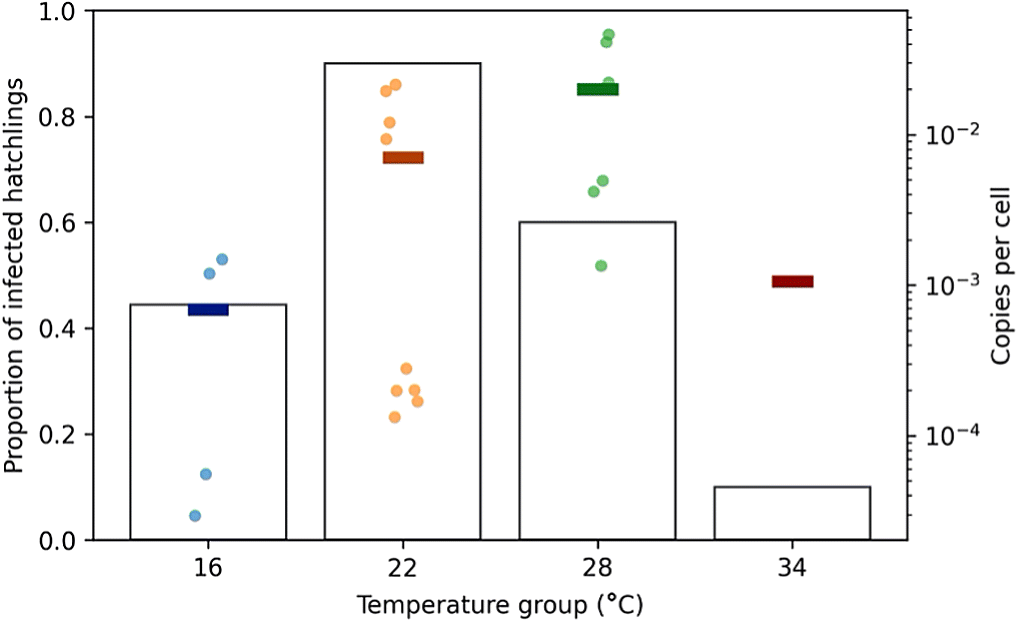
There was large variation in ranaviral MCP copy numbers within the different temperature groups (Fig. 1). As there was only one reactor in the 34 °C temperature group it was removed from the statistical analysis of viral loads. The Kruskal–Wallis test found a significant difference (0.05 α) between ranaviral MCP copy number between the 16, 22, and 28 °C temperature groups (H statistic = 6.57; p = 0.037). Post hoc analysis with the Conover’s test of multiple comparisons revealed that this difference was between the 16 and 28 °C temperature groups (p = 0.027).
Discussion
Based on the distribution of infection rates (Fig. 1) and temperature logger data (Table 2), the predicted optimal temperature for ranaviral infection (i.e., the maximum number of infected) in Krefft’s river turtles is 23.2 °C. Assuming that uninfected turtles would not later develop a detectable infection (i.e., turtles that did not react in the qPCR assay have cleared the infection), we can say that temperature has a significant effect on the rate and length of infection. This will have impacts on the transmission of the virus. The more animals infected and the longer an animal is infected, the greater the chance of viral transmission.
Clinical signs observed in this study were similar to those that were reported for ranaviral infection in other turtles, although not as extensive as previous reports (Wirth et al. 2019). Given more time these animals may have either recovered or developed more clinical signs; however, terminating the experiment at 21 d allowed us to prevent suffering of the animals (they did not have time to develop clinical signs) while still being able to study the effect of temperature on infection.
There was a large variation in the viral loads observed within temperature groups. Interestingly, the copies per cell of ranaviral DNA within each temperature group appears to divide equally into “high” and “low” reactors with at least a 10-fold difference between the mean high and low copy numbers. Such a difference in individuals that received the same treatment may be explained by some dichotomous biological trait, such as those associated with sex, that increases or decreases viral replication. Unfortunately, the sex of these hatchlings cannot be easily determined via physical exam and was not recorded at necropsy. Krefft’s river turtles do not have temperature-dependent sex determination, so the sex distributions within the temperature groups are assumed to be 50/50, which would explain the equal numbers of high and low viral loads in the temperature groups if this is the result of the sex of the animals. Future work should investigate the possible effects of sex (or other dichotomous variables) on ranaviral infection in turtles.
Post hoc analysis revealed that there was a significant difference in viral loads only between the 16 and 28 °C groups (p = 0.027). At lower temperatures, viral replication is likely reduced, thus resulting in reduced viral load. Adult red-eared sliders (Trachemys scripta elegans, suborder Cryptodira) exposed to a ranavirus and held at either 22 or 28 °C had significantly reduced ranaviral loads and halved morbidity in the 28 °C group (Allender et al. 2013). Similarly, we found that the infection rate was higher in the 22 °C group compared with the 28 °C group. However, we observed no significant difference in the viral loads between the 22 and 28 °C groups.
In a similar experiment, four Cryptodirid species of juvenile turtles (including red-eared sliders) did not have reduced mortality in the higher temperature group, both groups (22 and 27 °C) had 100% mortality (Allender et al. 2018). Krefft’s river turtles exhibit age-dependent ranaviral pathogenesis, with hatchlings being more susceptible than adult turtles (Ariel et al. 2015). The same may be true for red-eared sliders; the dose given to the juvenile turtles may have overwhelmed the turtles and thus negated the effect of temperature (Allender et al. 2013, 2018). Because adult red-eared sliders are less susceptible, the effect of temperature on viral replication/the immune system is still prominent (Ariel et al. 2015; Allender et al. 2018; Wirth et al. 2018).
In our study, we chose to use a median infectious dose (ID50), as determined by Wirth et al. (2019) for Krefft’s turtle hatchlings at 28 °C. As expected, the infection rate in the 28 °C group in this study was close to 50% (60%; Fig. 1). Using a median dose allows for easy detection of a change in the dependent variable (infection, death, etc.) as a result of changing different factors (e.g., environmental temperature). We chose the ID50 over the median lethal dose (LD50; 104.43 TCID50 mL−1) for this study for welfare reasons. The results of an infectious dose study are still valuable, although more applicable to a disease transmission than pathogenesis. We are not sure how these temperatures will affect Krefft’s river turtles exposed to a higher dose (such as an LD50). It may be that when Krefft’s turtle hatchlings are exposed to higher doses, the amount of virus will overwhelm any prophylactic effects of temperature, as seen with red eared-sliders. In any case, we still do not understand the natural transmissions dynamics, so it is difficult to determine what dose would be useful to accurately reflect wild disease.
Temperature therapy >34 °C may also be useful for treating acute ranaviral disease, although further study will be required to determine the effects of temperature on pathogenesis. While 34 °C appears effective at reducing infection rates, BIV itself is not inactivated at 34 °C (La Fauce et al. 2012); thus, once an animal returns to cooler temperature the infection may re-establish if the virus is not completely cleared. To reduce the chances of ranaviral infection in captive animals, we would recommend that turtles are provided with a basking area of at least 34 °C, thus allowing the animals to perform behavioural fever at temperatures high enough to reduce infection. Lower temperatures (e.g., ≤16 °C) may also be effective at reducing infection rates; however, these temperatures are not optimal for turtle health and result in reduced appetite and activity.
The optimal temperature for ranavirus isolate propagation in a range of cell lines is 24 °C (10, 15, 20, 24, and 28 °C tested; Ariel et al. 2009). The average annual temperature in Townsville (where this study was conducted) is 24.1 °C. Both the optimal temperature for ranaviral propagation and the average annual temperature in Townsville are close to the temperature of maximum infection rate (23.2 °C) estimated in this study. Australian freshwater turtles (like Krefft’s turtles) are semi-aquatic and spend the majority of their time in the water. When in the water, the turtles’ body temperatures are in thermal equilibrium with water temperature (Manning and Grigg 1997). Ranaviral infection rates would be expected to be highest in months when water temperature is around 23 °C, i.e., May–June and September–October. However, the temperature most effective at reducing infection rate used in this study (34 °C) is not uncommon in Northern Australia where these turtles are native. Thus, ranaviral-infected Krefft’s river turtles could have the opportunity to use behavioural fever to reduce infection rates, although finer-scale studies of thermal and basking site availability are required to quantify this opportunity.
In this study we, used reaction in a qPCR assay of the liver as a proxy for infection. While the liver is a major target of ranaviral replication, all conclusions reported in this study should be interpreted in light of the fact that reaction in a qPCR assay does not equal infectious virus.
Conclusions
Temperature is integral to ranaviral disease. We have shown that Krefft’s river turtles (suborder Pleurodira) exhibit temperature-dependent infection. Using the results of this study we have provided recommendations for temperature therapy of acute ranaviral infection, optimal temperature for behavioural fever in captive turtle enclosures and estimates of times of the year when infections rates are expected to be high in natural settings. Understanding how temperature influences emerging infectious diseases, like ranaviruses, is important in our changing climate. Results from studies like this one can provide data to climate change models and help form a foundation for understanding the impact on reptile populations.
Acknowledgements
The authors would like to acknowledge Anna Tzamouzaki for her help with husbandry, data collection, and sample processing; Bethany Adomanis for helping with husbandry; Lexie Edwards for helping with the experimental setup; and Lin Schwarzkopf for providing lab space, encouragement, and feedback.
References
Allender MC, Mitchell MA, Torres T, Sekowska J, and Driskell EA. 2013. Pathogenicity of frog virus 3-like virus in red-eared slider turtles (Trachemys scripta elegans) at two environmental temperatures. Journal of Comparative Pathology, 149: 356–367.
Allender MC, Barthel AC, Rayl JM, and Terio KA. 2018. Experimental transmission of frog virus 3–like ranavirus in juvenile chelonians at two temperatures. Journal of Wildlife Diseases, 54: 716–725.
Ariel E, and Jensen B. 2009. Challenge studies of European stocks of redfin perch, Perca fluviatilis L., and rainbow trout, Oncorhynchus mykiss (Walbaum), with epizootic haematopoietic necrosis virus. Journal of Fish Diseases, 32: 1017–1025.
Ariel E, Nicolajsen N, Christophersen M-B, Holopainen R, Tapiovaara H, and Jensen B. 2009. Propagation and isolation of ranaviruses in cell culture. Aquaculture, 294: 159–164.
Ariel E, Wirth W, Burgess G, Scott J, and Owens L. 2015. Pathogenicity in six Australian reptile species following experimental inoculation with Bohle iridovirus. Diseases of Aquatic Organisms, 115: 203–212.
Brand MD, Hill RD, Brenes R, Chaney JC, Wilkes RP, Grayfer L, et al. 2016. Water temperature affects susceptibility to ranavirus. EcoHealth, 13: 350–359.
Brunner JL, Storfer A, Gray MJ, and Hoverman JT. 2015. Ranavirus ecology and evolution: from epidemiology to extinction. In Ranaviruses. Edited by MJ Gray and VG Chinchar. Springer International Publishing, Cham, Switzerland. pp. 71–104.
Chinchar VG, Hyatt A, Miyazaki T, and Williams T. 2009. Family Iridoviridae: poor viral relations no longer. Current Topics in Microbiology and Immunology, 328: 123–170.
Chinchar VG, Hick P, Jancovich J, Subramaniam K, Waltzek T, Whittington R, et al. 2018. Create eight new species, remove three existing species in the family Iridoviridae. 2018.007D. [email protected] [online]: Available from talk.ictvonline.org/ictv/proposals/2018.007D.A.v1.Iridoviridae_8sp3sprem.zip.
Conroy CJ, Papenfuss T, Parker J, and Hahn NE. 2009. Use of tricaine methanesulfonate (MS222) for euthanasia of reptiles. Journal of the American Association for Laboratory Animal Science, 48: 28–32.
do Amaral JPS, Marvin GA, and Hutchison VH. 2002. The influence of bacterial lipopolysaccharide on the thermoregulation of the box turtle Terrapene carolina. Physiological and Biochemical Zoology, 75: 273–282.
Greenspan SE, Morris W, Warburton R, Edwards L, Duffy R, Pike DA, et al. 2016. Low-cost fluctuating-temperature chamber for experimental ecology. Methods in Ecology and Evolution, 7: 1567–1574.
Hall EM, Goldberg CS, Brunner JL, and Crespi EJ. 2018. Seasonal dynamics and potential drivers of ranavirus epidemics in wood frog populations. Oecologia, 188: 1253–1262.
Jensen BB, Holopainen R, Tapiovaara H, and Ariel E. 2011. Susceptibility of pike-perch Sander lucioperca to a panel of ranavirus isolates. Aquaculture, 313: 24–30.
La Fauce K, Ariel E, Munns S, Rush C, and Owens L. 2012. Influence of temperature and exposure time on the infectivity of Bohle iridovirus, a ranavirus. Aquaculture, 354–355: 64–67.
Leung WTM, Thomas-Walters L, Garner TWJ, Balloux F, Durrant C, and Price SJ. 2017. A quantitative-PCR based method to estimate ranavirus viral load following normalisation by reference to an ultraconserved vertebrate target. Journal of Virological Methods, 249: 147–155.
Maclaine A, Mashkour N, Scott J, and Ariel E. 2018. Susceptibility of eastern water dragons Intellagama lesueurii lesueurii to Bohle iridovirus. Diseases of Aquatic Organisms, 127: 97–105.
Manning B, and Grigg G. 1997. Basking is not of thermoregulatory significance in the “basking” freshwater turtle Emydura signata. Copeia, 1997: 579–584.
McKinney W. 2010. Data structures for statistical computing in Python. In Proceedings of the 9th Python in Science Conference (SCIPY 2010), 28 June–3 July 2010. Austin, Texas. 1697900, pp. 51–56.
Merchant M, Williams S, Trosclair PL III, Elsey RM, and Mills K. 2007. Febrile response to infection in the American alligator (Alligator mississippiensis). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 148: 921–925.
Monagas WR, and Gatten RE. 1983. Behavioural fever in the turtles Terrapene carolina and Chrysemys picta. Journal of Thermal Biology, 8: 285–288.
Reed LJ, and Muench H. 1938. A simple method of estimating fifty per cent endpoints. American Journal of Epidemiology, 27: 493–497.
Rojas S, Richards K, Jancovich JK, and Davidson EW. 2005. Influence of temperature on Ranavirus infection in larval salamanders Ambystoma tigrinum. Diseases of Aquatic Organisms, 63: 95–100.
Sauer EL, Trejo N, Hoverman JT, and Rohr JR. 2019. Behavioural fever reduces ranaviral infection in toads. Functional Ecology, 33: 2172–2179.
Seabold S, and Perktold J. 2010. Statsmodels: econometric and statistical modeling with Python. In Proceedings of the 9th Python in Science Conference (SCIPY 2010), 28 June–3 July 2010. Austin, Texas.
Spellerberg IF. 1972. Temperature tolerances of Southeast Australian reptiles examined in relation to reptile thermoregulatory behaviour and distribution. Oecologia, 9: 23–46.
Whittington RJ, and Reddacliff GL. 1995. Influence of environmental temperature on experimental infection of redfin perch (Perca fluviatilis) and rainbow trout (Oncorhynchus mykiss) with epizootic haematopoietic necrosis virus, an Australian iridovirus. Australian Veterinary Journal, 72: 421–424.
Wirth W, Schwarzkopf L, Skerratt LF, and Ariel E. 2018. Ranaviruses and reptiles. PeerJ, 6: e6083.
Wirth W, Schwarzkopf L, Skerratt LF, Tzamouzaki A, and Ariel E. 2019. Dose-dependent morbidity of freshwater turtle hatchlings, Emydura macquarii krefftii, inoculated with Ranavirus isolate (Bohle iridovirus, Iridoviridae). Journal of General Virology, 100: 1431–1441.
Zimmerman LM, Vogel LA, and Bowden RM. 2010. Understanding the vertebrate immune system: insights from the reptilian perspective. Journal of Experimental Biology, 213: 661–671.
Information & Authors
Information
Published In

FACETS
Volume 5 • Number 1 • January 2020
Pages: 821 - 830
Editor: David Lesbarrères
History
Received: 28 February 2020
Accepted: 19 May 2020
Version of record online: 22 October 2020
Notes
This paper is part of a Collection titled “Ranavirus research: 10 years of global collaboration.”
Copyright
© 2020 Wirth and Ariel. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper.
Key Words
Sections
Subjects
Authors
Author Contributions
All conceived and designed the study.
WW performed the experiments/collected the data.
WW analyzed and interpreted the data.
All contributed resources.
All drafted or revised the manuscript.
Competing Interests
Ellen Ariel is a guest editor.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Wytamma Wirth and Ellen Ariel. 2020. Temperature-dependent infection of freshwater turtle hatchlings, Emydura macquarii krefftii, inoculated with a ranavirus isolate (Bohle iridovirus, Iridoviridae). FACETS.
5(1): 821-830. https://doi.org/10.1139/facets-2020-0012
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Low-temperature immunization attenuates the residual virulence of
orf074r
gene-deleted infectious spleen and kidney necrosis virus: a candidate immersion vaccine
2. Trace Amounts of Ranavirus Detected in Common Musk Turtles (Sternotherus odoratus) at a Site Where the Pathogen Was Previously Common
3. Nocturnal basking in freshwater turtles: A global assessment
4. A review of reptile virus experimental infection studies
5. Pathogenesis of Bohle iridovirus infection in Krefft’s freshwater turtle hatchlings (
Emydura macquarii krefftii
)