Introduction
Evidence for the effect of turbine size on bird and bat fatalities is equivocal (
Table 1), possibly reflecting the wide variation in locations, affected species, and turbine type and sizes among studies. Variation in the area searched under turbines also affects fatality estimates. Taller turbines propel carcasses further, potentially causing them to land outside standard search areas (
Hull and Muir 2010;
Huso and Dalthorp 2014;
Choi et al. 2020) and biasing estimates of fatalities low at taller turbines compared to shorter turbines with equivalent search areas. Finally, low sample sizes can preclude analyses of species-specific patterns in fatalities in studies based on smaller datasets, but analyzing total fatalities (i.e., with all species or groups of species pooled) may obscure patterns between turbine height and species-specific fatalities (
Davy et al. 2020).
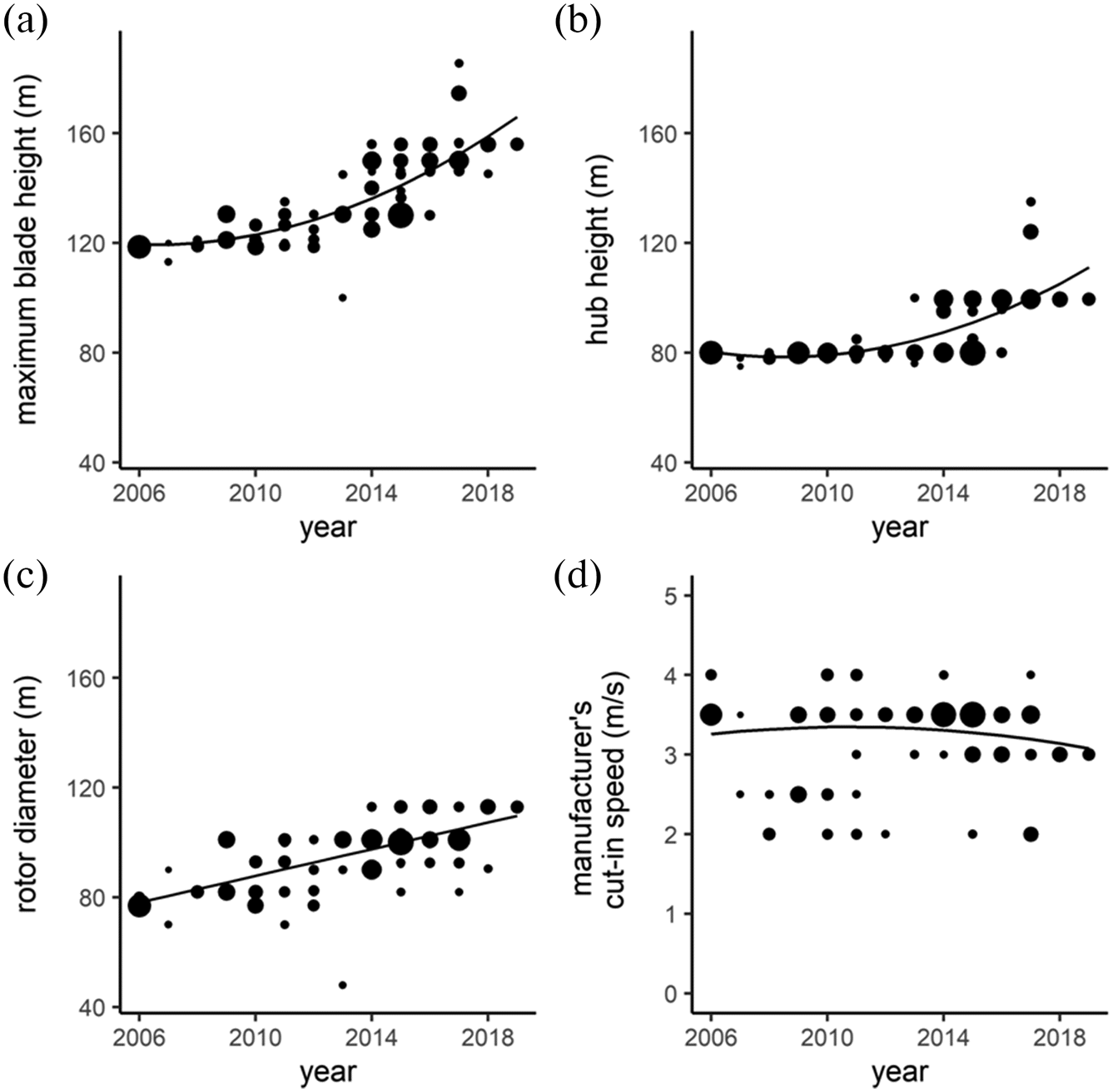
Wind turbines in Ontario, Canada, have increased 42% in size, from 120 to 170 m maximum blade height (MBH), between 2006 and 2019 (
Fig. 1a and
Table S1), a result of increases in hub height (
Fig. 1b) and rotor diameter (
Fig. 1c). A previous analysis suggested that turbine hub height may contribute to fatalities of some bat species in Ontario, but confidence in the relationship was not strong (
Davy et al. 2020). Here, we conducted a detailed investigation of the effects of turbine height on fatalities in two groups of aerial insectivores, bats (nocturnal) and swallows (diurnal), which are experiencing population declines (
Nebel et al. 2010;
Committee on the Status of Endangered Wildlife in Canada 2013;
Smith et al. 2015;
Rodhouse et al. 2019). We expanded on previous work by including data from newer, taller turbines and accounting for carcasses falling outside of search areas at taller turbines. We also used MBH as a measure of turbine height because it is more relevant than hub height to measure the altitudes where species may interact with turbine blades.
Methods
Data sources
We obtained searcher efficiency, scavenging, and carcass survey data from post-construction monitoring of onshore monopole turbines at wind energy facilities in southern Ontario (south of 48°N) and submitted to the Wind Energy Bird and Bat Monitoring Database (
Birds Canada 2020). The database included turbine model names, hub heights, and rotor diameters gathered from facility reports or online (
thewindpower.net) and manufacturer-specified cut-in speeds from a repository of wind turbine models (
wind-turbine-models.com/turbines).
We used MBH (hub height plus half the rotor diameter) to quantify turbine height in our analyses because hub height and rotor diameter are correlated, and MBH is more ecologically meaningful than hub height alone. Hub height and rotor diameter are moderately correlated in Ontario (
ρ = 0.49), and including both variables in models could lead to unstable parameter estimates, inflated standard errors, and difficulty assessing the relative importance of the variables (
Dormann et al. 2012). Additionally, MBH incorporates hub height and rotor diameter into a single metric representing the true height of a turbine, which has implications for animals flying at different altitudes.
Study species
We modelled the relationship between MBH and fatalities of each bat and swallow species sufficiently represented in the database. These included five species of bat (big brown bat Eptesicus fuscus Palisot de Beauvois, 1796; little brown bat Myotis lucifugus Le Conte, 1831; eastern red bat Lasiurus borealis Müller, 1776; hoary bat Lasiurus cinereus Palisot de Beauvois, 1796; and silver-haired bat Lasionycteris noctivagans Le Conte, 1831) and four species of swallow (barn swallow Hirundo rustica Linnaeus, 1758; tree swallow Tachycineta bicolor Vieillot, 1808; cliff swallow Petrochelidon pyrrhonota Vieillot, 1817; and purple martin Progne subis Linnaeus, 1758).
Fatality estimation
We estimated the number of individuals of each species killed (
) at monitored wind turbines using GenEst (
Dalthorp et al. 2018) in R (
R Core Team 2020). GenEst is a generalized estimator that corrects the number of carcasses found by the probability of detecting a carcass and adjusts for carcasses falling outside of searched areas. Details of the specifications we used to calculate
in GenEst follows.
Search schedules
We retained data from wind turbines that were consistently monitored (twice per week for 98% of datasets, 2% searched once per week) during spring (May–Jun) and (or) late summer (mid-Jul through end-Sep). We only estimated fatalities for swallows in late summer because few carcasses were found at turbines during spring (n = 19, 6% of all swallow carcasses found in our data subset).
GenEst requires users to enter search dates for each turbine to estimate mortality, including searches where no carcasses were found. However, only search dates when carcasses were found were available for a subset of facilities and years (
n = 28, 23% of all facility years). To create full search schedules for these facilities and years, we simulated probable search dates for each turbine using rule-based data mining and hierarchical clustering analysis in the
arules package in R (
Hahsler et al. 2005,
2020). In short, we used a priori association rules to group turbines that were often searched on the same days and assigned search dates to the whole group. We then filled gaps in the search schedules of each group so that searches occurred at intervals specified in the database (e.g., every 3 or 4 days for twice-weekly searches) and throughout the survey period (e.g., 15 Jul–30 Sep for late summer).
Detection probability
We used facility- and year-specific searcher efficiency and carcass persistence survey data to produce facility-corrected fatality estimates. Our analysis included facilities and years for which ≥10 searcher efficiency and ≥10 carcass persistence surveys were conducted (
n = 59 facilities surveyed in at least one year during 2010–2019). Carcasses used in searcher efficiency and carcass persistence surveys were small bats and non-raptor birds. We assumed a constant proportional decline in searcher efficiency (
k = 0.7) with each subsequent search (
Korner-Nievergelt et al. 2012;
Simonis et al. 2018), because carcasses missed on the first search may be more difficult to find as a result of decay, cryptic colouration, etc. We could not estimate
k from available data because test carcasses were removed after each searcher efficiency trial. We modelled carcass persistence over time with exponential, Weibull, lognormal, and loglogistic survival models and constant scale and location parameters. Of the four candidate models for each facility and year, we selected the top model with the lowest ΔAICc value.
Search area correction
All carcass searches were conducted by walking concentric circles out to a maximum distance of 50 m (as required by government guidelines;
Ontario Ministry of Natural Resources and Forestry 2011), regardless of turbine height. Ballistics models estimate that ∼95% of 14 g bat carcasses (and ∼80% of 12 g bird carcasses) fall within 50 m of turbines with MBH = 125 m (i.e., “medium-sized” turbines;
Hull & Muir 2010), which were typical sizes of turbines on the landscape at the beginning of our study period (2010,
Fig. 1a). However, newer turbines are now much larger (∼170 m max turbine blade height in 2019,
Fig. 1a), and therefore, a larger proportion of carcasses should land outside of 50 m search areas than for older, shorter turbines.
We used ballistics models (
Hull and Muir 2010), field observations of carcasses, and the proportion of the search area covered by searchers to estimate the proportion of carcasses expected to fall in the search area of each turbine (
dwp). First, for each type of turbine at a facility, we estimated the maximum distance that bat and small bird carcasses should fall from the base of turbines (henceforth, “maximum fall distance”) depending on the blade length (half of rotor diameter) and hub height (i.e., from linear regression equations,
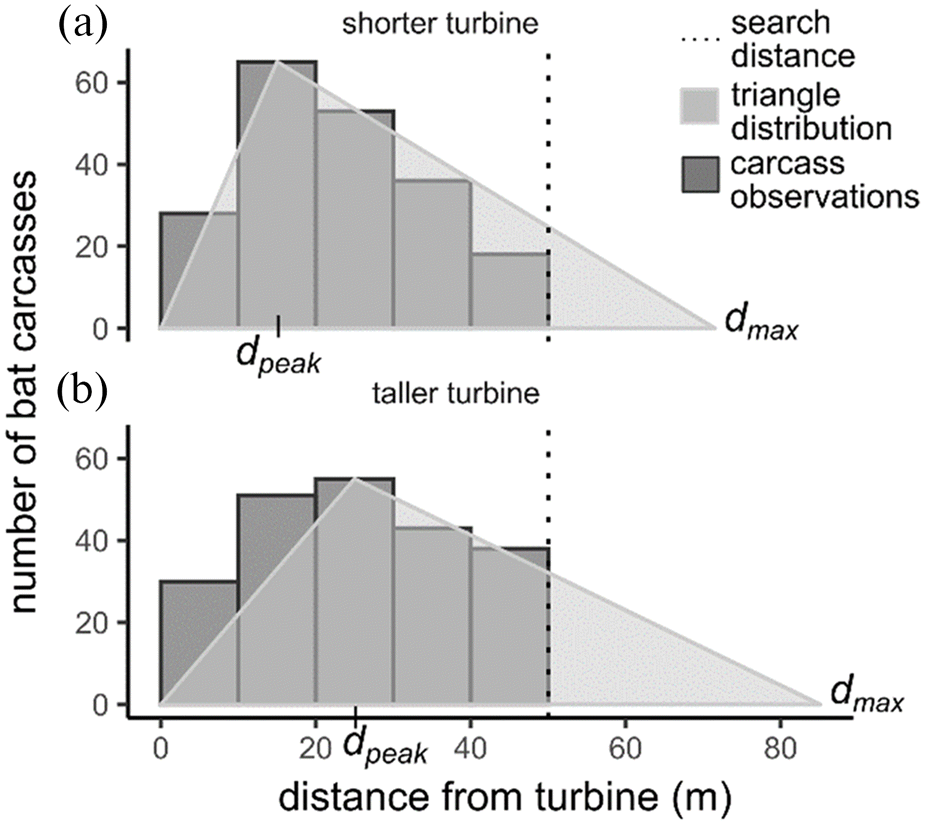
Hull and Muir 2010). Next, for the same turbine groups, we counted the number of bat and bird carcasses found every 10 m from the base of turbines out to 50 m. Carcasses tended to peak closer to the turbine base at shorter turbines (
Fig. S1) and roughly followed a triangular distribution (
Fig. 2). We assumed that the triangular distribution extended to the maximum fall distance, with the number of carcasses declining linearly from the peak out to the maximum fall distance. We then estimated the proportion of carcasses that should fall in the 50 m search radius (
psearch) as the proportion of the total triangular distribution within 50 m of the turbine base (
Fig. 2). Carcass distribution data were limited or unavailable for a subset of facilities (23% of facilities for bats and 49% for birds), so we estimated
psearch for the subset using predictive equations from a linear regression between calculated
psearch estimates and taxa type (bird or bat) and MBH (
Fig. S2). Finally, we estimated
dwp for each turbine by multiplying
psearch for each facility and turbine type by the average percent of the 50 m search area covered by searchers.
Estimation
We used search schedule,
dwp, carcass persistence, searcher efficiency, and carcass observation data to calculate
using the estM function with 10 000 iterations and the calcSplits function to estimate
per turbine and season (spring or late summer). We used the R package
purrr (
Henry and Wickham 2020) to simultaneously estimate
for each facility, year, and species, and we retained median
values for analyses in linear mixed effects models.
Mixed effects models
We examined the effects of MBH on bat and swallow fatalities using generalized linear mixed effects models (GLMMs) with per turbine as the response variable. We included species and manufacturer’s cut-in speed as predictors, as well as the interactions between species and MBH, and species and cut-in speed. In addition, we included turbine ID and facility ID as random factors to account for repeated measures and for other potential turbine- or facility-specific sources of variation. We could not include year as a continuous predictor because the number of facilities monitored in some years was low (n = 2 facilities in 2010, 2012, and 2019, respectively). We, therefore, included a random factor of year to estimate the effects of turbine height and cut-in speed while accounting for annual variation in fatalities.
We fit GLMMs and zero-inflated GLMMs with Poisson and negative binomial distributions using the R package
glmmTMB and compared the fits of global models using AIC (
Brooks et al. 2017) via the function “AICtab” in the
bbmle package (
Bolker and R Development Core Team 2020). We also used the package
DHARMa (
Hartig 2020) to examine patterns in residuals. We selected the global model with the best fit, indicated by the lowest ΔAIC: zero-inflated negative binomial models for bats and swallows in late summer and zero-inflated Poisson models for bats in spring (
Table S2). We subsequently used “drop1” function with a chi-square test to drop non-significant terms (
α = 0.05) from the model. We calculated marginal and conditional
R2 following
Nakagawa et al. (2017).
Discussion
We estimated fewer bat fatalities at turbines undergoing operational mitigation (i.e., raising cut-in speeds), which agrees with previous findings (
Arnett et al. 2011;
Davy et al. 2020;
Whitby et al. 2021). Despite fewer fatalities at turbines with raised cut-in speeds, cut-in speeds specified by manufacturers were poor predictors of estimated bat and swallow fatalities. Bat activity tends to drop sharply at wind speeds greater than 5–6 m/s (
Reynolds 2006;
Wellig et al. 2018); therefore, bat activity may not have differed substantially between wind speeds of 2–4 m/s (the range of cut-in speeds specified by manufacturers in our dataset). However, turbine-specific operational cut-in speeds from our study area were not available, so we used manufacturer’s specified cut-in speeds as a proxy for operational cut-in speeds. We recognize that wind farms sometimes allow turbines to “free-wheel” (i.e., to continue to spin at low wind speeds prior to the onset of electricity generation) and that some facilities may have used different cut-in speeds than that specified by the manufacturer, and both scenarios would obscure the relationship we predicted. Wildlife collide with both free-wheeling turbines and turbines that are generating power (
Baerwald et al. 2009;
Whitby et al. 2021), so if turbines in our study area were free-wheeling, then manufacturer specified cut-in speed would become a poor proxy for turbine movement at low wind speeds and a poor predictor of fatalities. To address this potential confounding factor, we recommend that future studies obtain and examine operational cut-in speeds and data on free-wheeling turbines whenever possible.
Population size and flight altitudes of bats are not well understood (
Voigt et al. 2018) relative to swallows’ population size and flight altitudes (
Table S4). Although few swallow carcasses (
n = 341) were found at turbines compared to bats (
n = 4850), we speculate that quantitative data about swallow flight altitudes, population size, and fatalities at turbines could provide biologically meaningful interpretations of bat fatality data. Despite their higher relative abundance in Ontario (
Partners in Flight 2020;
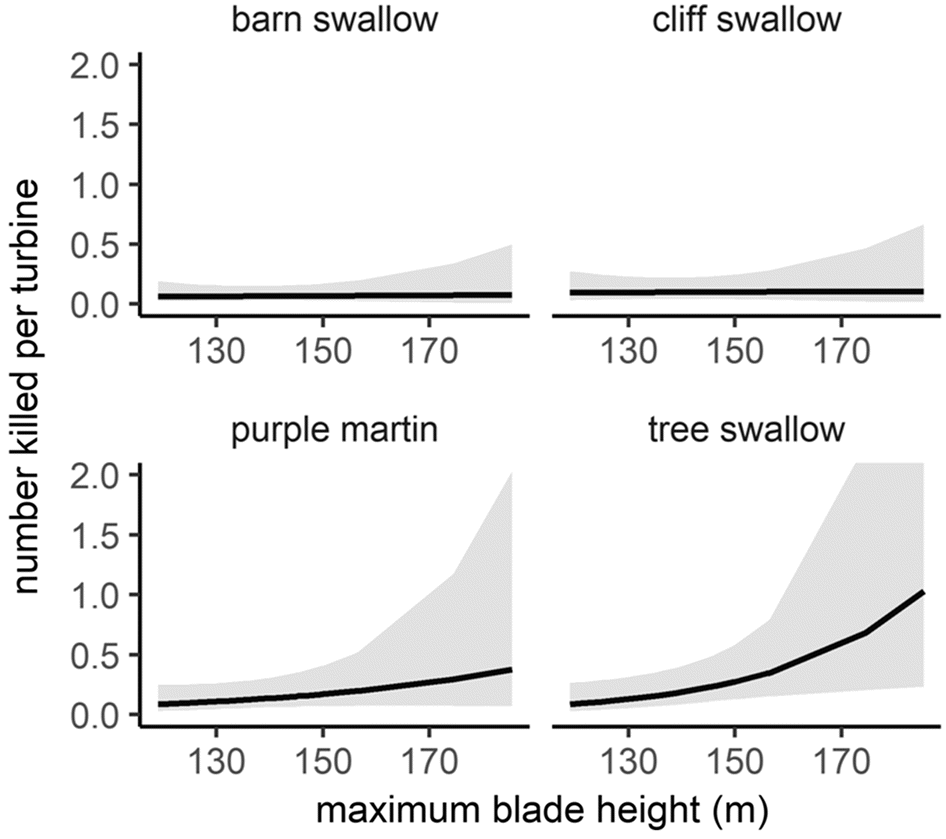
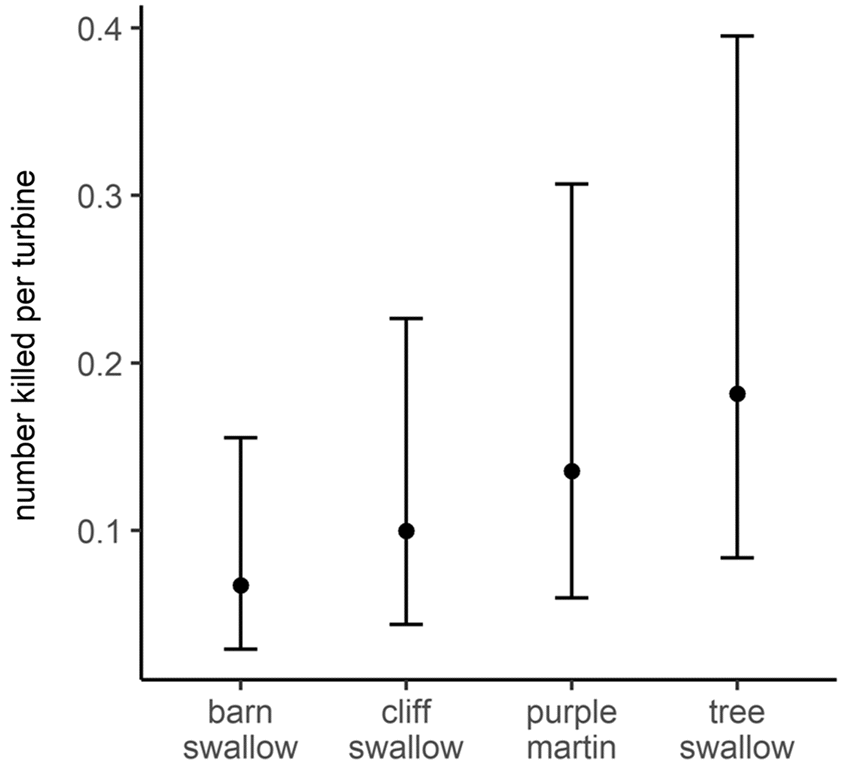
Table S4), barn swallows had the lowest per turbine fatalities compared to other higher-flying swallow species (e.g., tree swallow and purple martin;
Dreelin et al. 2018). Additionally, fatalities of purple martins, the least abundant but highest-flying species in Ontario (
Table S4), were greater at taller turbines than shorter turbines. We, thus, speculate that differences in fatalities of avian aerial insectivores at turbines are due to differences in flight altitude, not necessarily abundance. Likely, both flight altitude and relative abundance contribute to fatalities, as demonstrated by tree swallow fatality data; tree swallows had the highest number of fatalities per turbine, and they are abundant (
Table S4) and also often fly in the rotor swept area of turbines in Ontario (
Fig. S3).
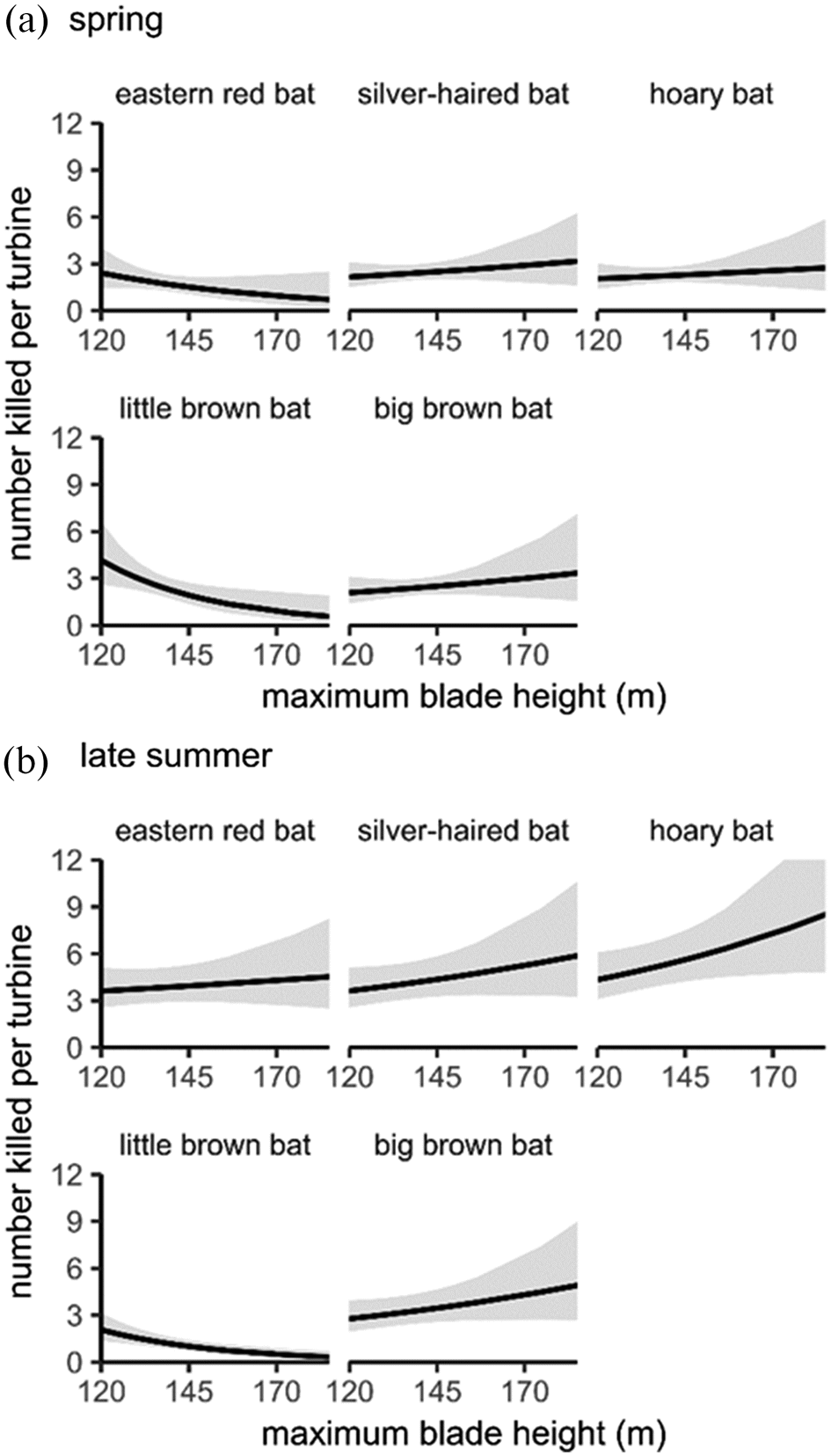
If the same relationship between flight altitude and fatalities holds true for bats, then the pattern we observed between turbine height and fatalities of hoary, silver-haired, and big brown bats may reflect these species use of higher altitudes. Collisions of hoary and silver-haired bats with planes support the assertion that they are high-altitude flyers (
Peurach 2003;
Peurach et al. 2009;
McCracken et al. 2021). Perhaps little brown bats and eastern red bats fly at lower altitudes, spending more time below the rotor swept area of newer turbines that tend to be further from the ground than older turbine models, but at this point, this interpretation is speculation that is not yet supported by quantitative data. New, lightweight tracking devices such as GPS units and altimeters can help explicitly test hypotheses about bat flight altitudes (
Roeleke et al. 2016;
Weller et al. 2016;
Conenna et al. 2019;
O’Mara et al. 2019,
2021; though see
Péron et al. 2020).
Broadly, our results and those of other studies (
Johnson et al. 2003;
Fiedler et al. 2007;
Barclay et al. 2007;
Baerwald and Barclay 2009;
Rydell et al. 2010;
Georgiakakis et al. 2012) indicate a general, ongoing pattern of more bat fatalities at taller turbines. While more bat fatalities at taller turbines is a concern with the increasing size of turbines (
Wiser et al. 2011,
2018), fatality rates of bats and birds appear to be relatively consistent per unit of energy produced (GWh) by a turbine (
Huso et al. 2021). Therefore, taller turbines that tend to produce more energy may kill more birds and bats than smaller turbines, but the number of animals killed per GWh may remain constant. If we consider how this result can inform efforts to maximize energy production while minimizing wildlife collisions, considering turbine height in the new construction or the repowering of turbines is likely a less effective strategy for reducing wildlife fatalities than efforts such as minimizing turbine blade movement during high-risk periods, for example at night, during migratory periods, and in certain weather conditions for bats (
Arnett et al. 2011;
Squires et al. 2021) and during the post-breeding and southbound migration period for swallows (
Erickson et al. 2014).
Nevertheless, our species-level analyses reveal that turbine height does predict the number of fatalities for some species of bats and birds. Where possible, future analyses of wildlife mortality at wind turbines should explicitly consider interspecific variation in fatalities at wind turbines. Species-specific analyses can inform more accurate risk assessments for each species and may provide insight into aerial habitat selection by each species. Finally, we acknowledge that using mortality as a proxy for exploring flight altitudes is indirect and laden with assumptions. We expect that the increasing availability of altimeters for wildlife tracking will improve interpretation of fatality data at wind turbines and clarify the association between flight altitude and collision risk.