Introduction
Seasonal monitoring of seagrass and adjacent estuarine benthic habitats improves understanding of changes in the meadow and the provision of ecosystem services but requires methods that can detect fine-scale seasonal variability in cover and extent of the different benthic habitats (
Chand and Bollard 2022), while surveying at the meadow-scale (often several hectares). These methods must also be reproducible, meaning they produce similar quality data across surveys. Numerous approaches have been used for seasonal monitoring of seagrass meadows including wading/snorkel/SCUBA surveys (
Cho et al. 2017;
Fourqurean et al. 2001;
Short et al. 2006), aerial photos (
Reise and Kohlus 2008), satellite imagery (
Lyons et al. 2013), and acoustic surveys (
Sonoki et al. 2016). Remote sensing applications for seagrass monitoring are generally used to measure seagrass extent (
Lyons et al. 2013), whereas wading/snorkel/SCUBA approaches reveal smaller scale and higher resolution details such as shifts in percent cover of seagrass, shoot density, and species composition (i.e., presence of macroalgae;
Fourqurean et al. 2001;
Short et al. 2006). More recently, remotely piloted aircraft systems (RPAS; i.e., drones) have been applied to monitor seasonal variation in seagrass extent and structure (
Chand and Bollard 2022;
Krause et al. 2021) due to their ability to collect high resolution images frequently and at a relatively low cost and their potential to automate part of the work (
Colefax et al. 2018;
Joyce et al. 2018). By reducing monitoring costs, RPAS present a promising approach for seagrass monitoring because seagrass monitoring is widespread and conducted by groups that are resource and time limited, including academics, non-profit, and government entities seeking to optimize monitoring resources. However, studies on seasonal reproducibility of RPAS did not include delineation of other adjacent estuarine habitats (e.g., other species of submerged aquatic vegetation), which can be challenging to spectrally separate (
Duffy et al. 2018;
Tait et al. 2019), and focused RPAS surveys on limited portions of a meadow. Examining whether full-meadow scale seasonal RPAS surveys can reliably detect changes in seagrass extent, percent cover, and differentiate seagrass from other habitats would determine whether RPAS effectively bridge the gap between remote sensing and wading/snorkel/SCUBA surveys.
To detect seasonal changes in seagrass, RPAS surveys must be reproducible. However, a major challenge with RPAS surveys is that image quality quickly diminishes in less than optimal environmental conditions (e.g., cloud cover, wind, turbidity, high-tide), challenging seagrass identification (
Joyce et al. 2018;
Nahirnick et al. 2019a,
2019b). These environmental changes can shift within minutes/hours and mid-survey (e.g., tide, cloud cover, sun angle) as well as over longer time periods, including seasonally and annually. Additionally, seagrass blades may fold over at low tide or when there is a fast current, increasing the likelihood of overestimating seagrass density. While recent studies have provided roadmaps of ideal environmental conditions for RPAS surveys of seagrass (e.g.,
Joyce et al. 2018;
Nahirnick et al. 2019b;
Tait et al. 2019;
Yang et al. 2020), studies evaluating the efficacy of surveys have generally focused on one-time occurrences in tropical regions (e.g.,
Ellis et al. 2020) or portions of a meadow (
Barrell and Grant 2015;
Duffy et al. 2018;
Ellis et al. 2020;
Konar and Iken 2018;
Krause et al. 2021). In temperate regions, high environmental variability, including rapid shifts in cloud cover, wind, and chemistry of estuarine waters (e.g., tannins), challenges RPAS surveys (
Nahirnick et al. 2019a,
2019b). The source and frequency of environmental variability in temperate estuarine systems often differ among seasons, making seasonal monitoring even more challenging as it can be difficult to collect comparable images. For instance, seasonal effects include higher rainfall in the fall and snowmelt in the spring, whereas daily variation includes changing water levels due to tide and rainfall and varying tannin concentrations depending on riverine discharge. Therefore, an evaluation of survey accuracy and reproducibility in such systems is necessary if RPAS are to be used to monitor changes in habitat extent, percent cover, and adjacent benthic habitat over time.
Detecting fine-scale seasonal changes in seagrass meadows requires collecting high spatial resolution surveys. RPAS image resolution is inversely proportional to altitude, and fewer details in images collected at higher altitudes can complicate habitat classification and survey reproducibility (
Seifert et al. 2019). However, increasing survey altitude reduces flight time as larger areas are covered per image, which limits the risk of environmental conditions such as sun angle and tide, which change within minutes/hours, from changing mid-survey (
Joyce et al. 2018) and enables more frequent surveys. Furthermore, varying flight altitude may affect the reproducibility of RPAS surveys in multiple ways.
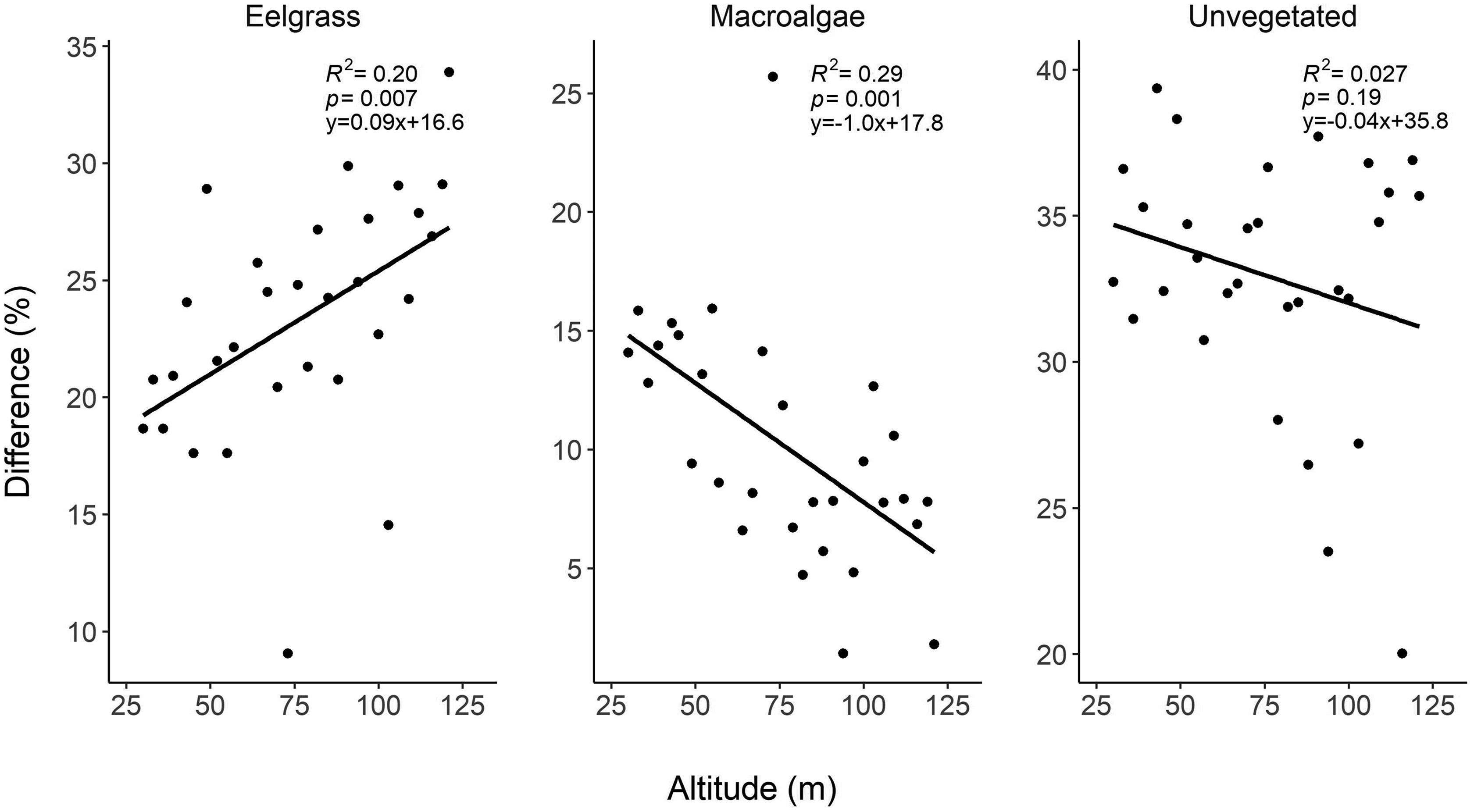
Ellis et al. (2020) showed the effect of altitude on image classification depended on habitat when surveying at increasing altitude, presumably due to different levels of spectral separability among habitats. Sand cover estimates increased with altitude, whereas seagrass estimates decreased, and mangrove, coral, and open water estimates were unaffected. These differences highlight the need to better understand the trade-offs associated with altitude, including how they relate to the structure of the meadow (e.g., fragmented vs. continuous) and location in the meadow (i.e., edge vs. middle) to conduct reproducible seasonal surveys of seagrass meadows.
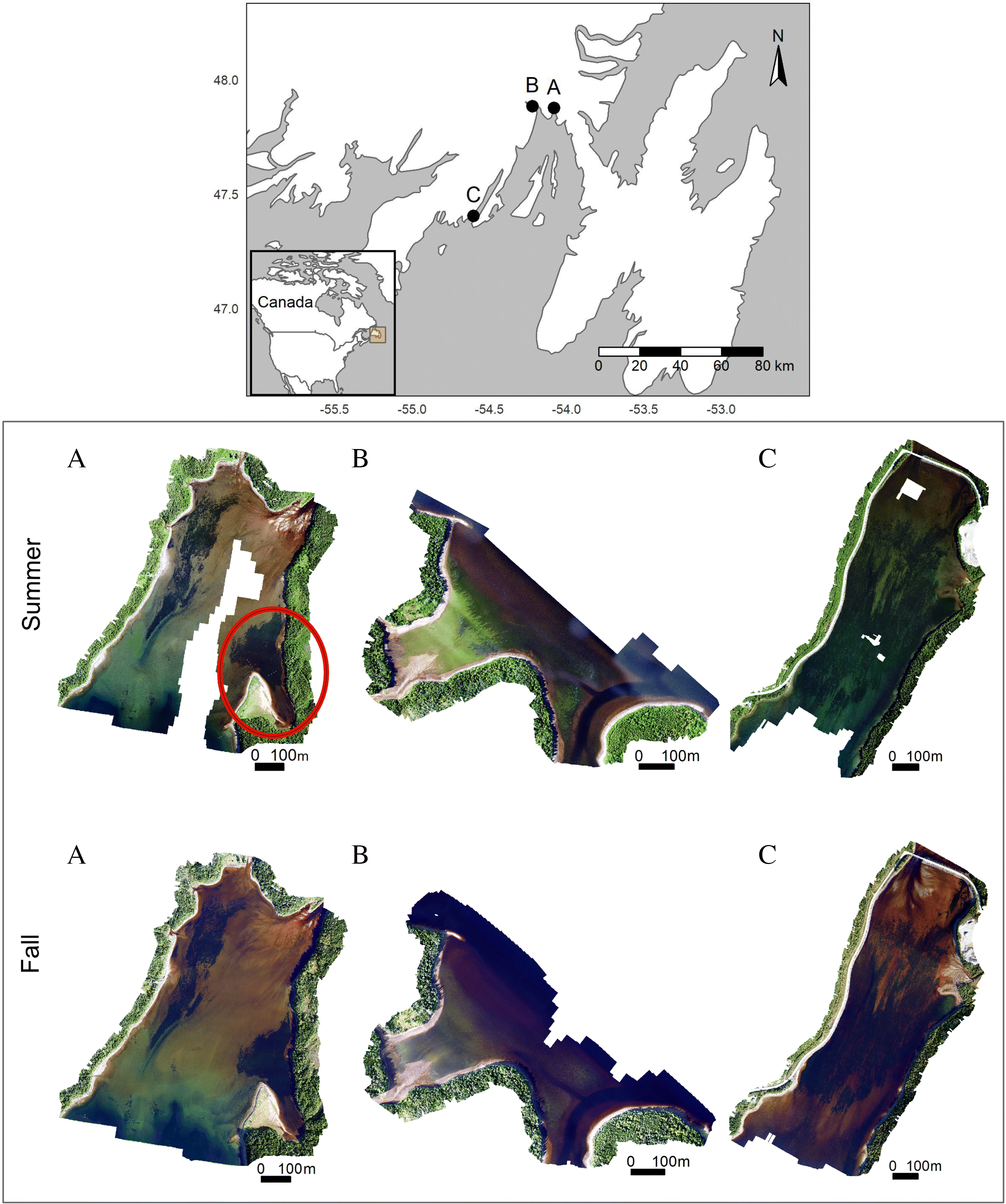
The first objective of this study was to test the reproducibility for RPAS to monitor seasonal changes in eelgrass (Zostera marina) meadows and adjacent benthic estuarine habitats in cold-temperate ecosystems. This represents the first study to evaluate the capacity of RPAS surveys to detect seasonal (i.e., summer vs. fall) variation in meadow extent, percent cover, and macroalgae cover across multiple sites of relatively large area (∼50 ha). We compared orthomosaics and image classification accuracies across surveys to assess the reproducibility of RPAS for estimating seasonal changes in seagrass extent in three sites. RPAS estimates of habitat percent cover and species composition were compared to complementary snorkel quadrat surveys to evaluate whether both methods yielded similar seasonal trends in eelgrass and macroalgae percent cover. Finally, since flight altitude implies trade-offs between image resolution and survey time, our second objective was to evaluate the effects of flying at a high altitude (e.g., 30 m vs. 115 m) on estimates of seasonal changes in percent cover.
Discussion
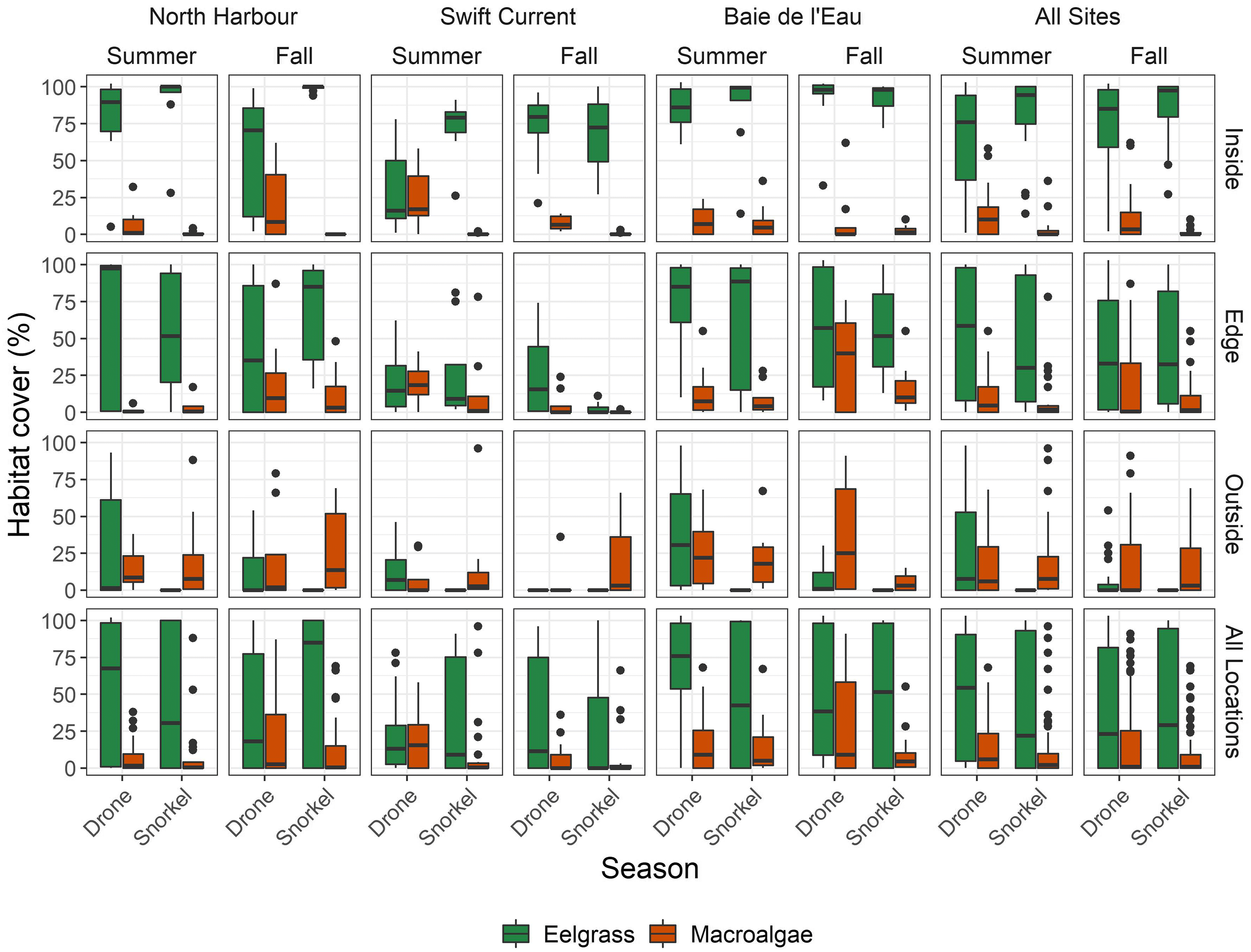
This study assessed the feasibility of conducting reproducible RPAS surveys to monitor the seasonal dynamics of eelgrass and adjacent benthic habitats at the meadow-scale (e.g., 50 ha) in a cold, temperate, estuarine ecosystem. Our results demonstrate that the capacity of RPAS to detect seasonal variation in habitat percent cover is highly variable within and among sites. The two survey methods (i.e., RPAS and snorkel transects) used in this study detected a <1% seasonal change in eelgrass percent cover, suggesting negligible variation in seagrass cover and therefore, potentially negligible change in the provision of ecosystem services by eelgrass in Newfoundland between mid-summer and early-fall. Eelgrass in Atlantic Canada reaches peak growth in August before declining in the winter due to sea ice and storms (
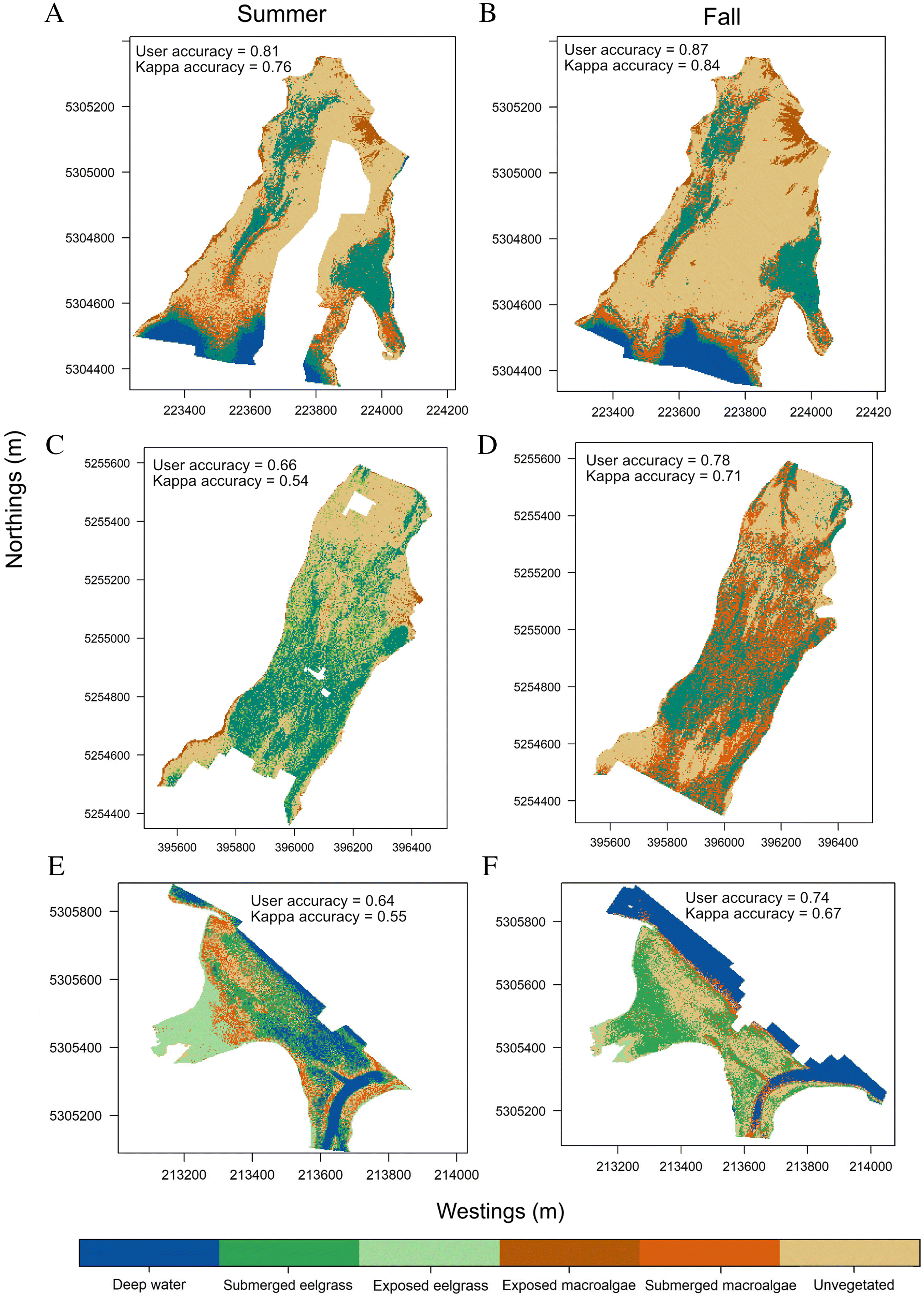
Murphy et al. 2021). Therefore, our study shows that annual monitoring of eelgrass during peak season could be extended until late September/early October. However, RPAS image classification was challenged by fragmented, heterospecific, and deeper benthic habitats. Inconsistent misclassification rendered locating micro/meso-scale changes infeasible, potentially preventing us from identifying sections of the meadow where seasonal changes in extent and percent cover may have occurred. This would have implications for the reproducibility of any repeated monitoring using RPAS, whether it be seasonal or annual.
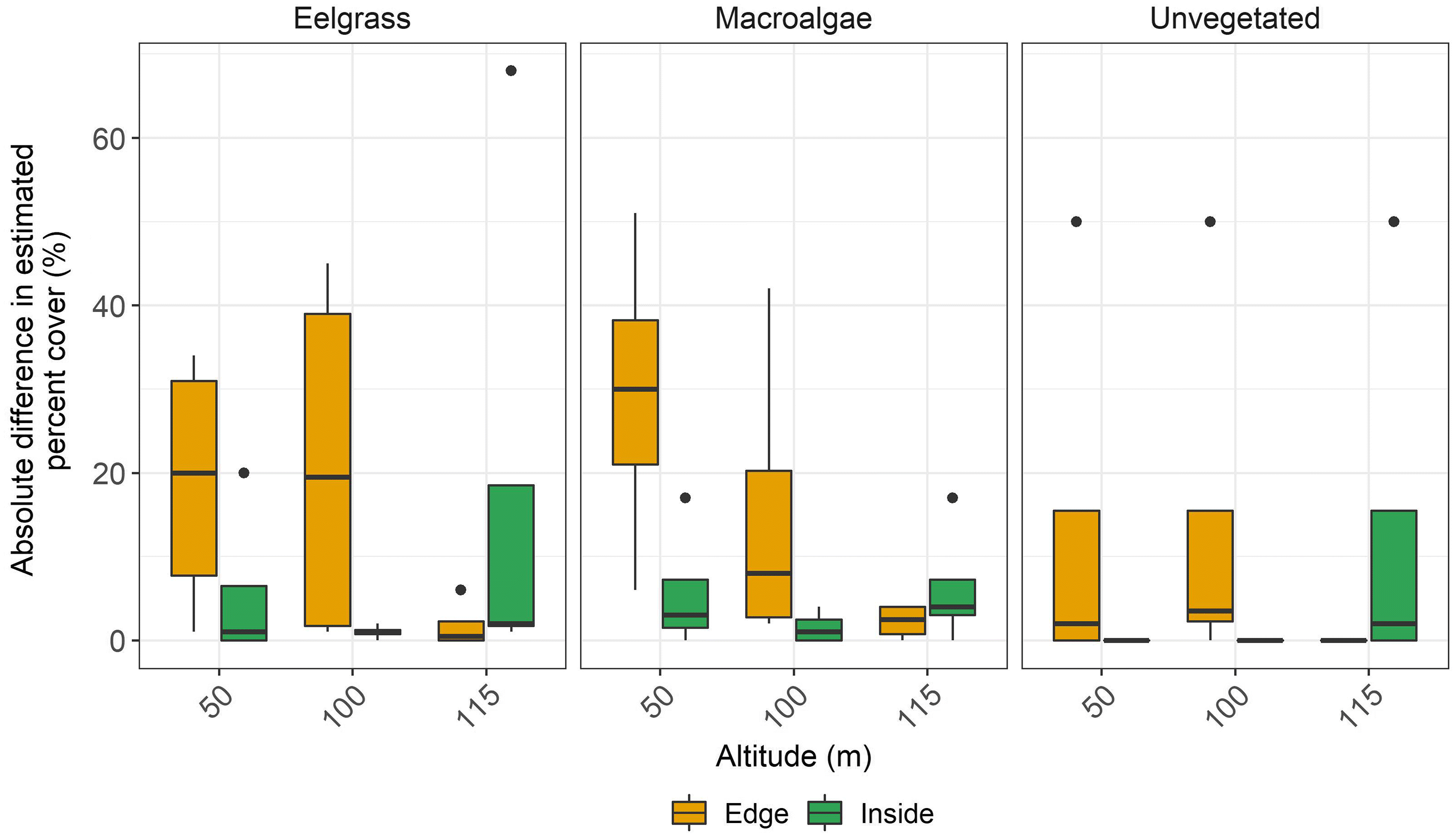
Our results also showed that the effects of flight altitude on accuracy are not straightforward. Macro-scale (i.e., snorkel transect comparison) estimates of cover improved with increased altitude, whereas micro/meso-scale (i.e., submerged quadrat comparison) cover estimates varied depending on the habitat. Flying at higher altitude advantageously reduces survey time to the detriment of image resolution. However, given that accuracy was not necessarily reduced at higher altitude, we suggest that, when monitoring macro-scale changes in benthic estuarine habitats where the environmental conditions are frequently shifting within minutes/hours, the benefit of increased likelihood of successful survey (e.g., image stitching) outweighs the cost of collecting lower resolution imagery. This finding is applicable to any group seeking to use RPAS to monitor macro-scale changes in submerged, estuarine habitats.
Reproducibility
The reproducibility of RPAS surveys is likely limited by a combination of environmental factors. Successfully mapping eelgrass meadows using RPAS has been linked to weather and light penetrability (
Joyce et al. 2018;
Nahirnick et al. 2019a,
2019b). Although all surveys in our study were conducted in optimal weather conditions and all our sites were less than 5 m deep, depth was not consistent among surveys as we could not control for tide. Light penetration is reduced with increasing depth, such as high tide or deeper portions of a meadow. This is particularly the case for red wavelengths, which become absorbed or scattered, making deeper sites harder to survey (
Duntley 1963;
Tait et al. 2019). In shallow portion of the meadow, low tide causes seagrass blades to bend, which increases the likelihood of overestimating seagrass percent cover and extent. As a result, there is no universal ideal tidal height to survey seagrass using RPAS. The effect of tide on seagrass imagery varies depending on the meadow blade length and how much depth varies throughout the meadow. We did not have the data resolution to detect whether water tidal height caused the variability we observed; however, neither did we observe any patterns in our results.
Turbidity, a common feature in coastal temperate regions due to freshwater influx and seasonal phytoplankton blooms, also inhibits light penetrability and is a common feature in coastal temperate regions (
Babin et al. 2003;
Duntley 1963). Turbidity was low for all surveys (
Table 2) and therefore was unlikely to have driven the differences in accuracy. However, we did observe seasonal differences in the surface water color, especially in the Baie de l’Eau fall survey (
Fig. 1C). This was likely caused by influx of tannin-rich freshwater, as observed by the red/brown color of the orthomosaic (
Fig. 1C), and potentially by the tidal amplitude difference between surveys (1.4 m compared to 0.2 m for the other two sites). Seasonal variation in freshwater influx and mixing generates unique interactions among light, depth, and color. These interactions may have contributed to the seasonal variability in survey accuracy observed in our study.
As observed in other studies, spectral separability between habitats affected image classification (
Lillesand and Kiefer 2000;
O’Neill and Costa 2013;
Tait et al. 2019;
Ventura et al. 2018). This in turn affected the reproducibility of our RPAS eelgrass surveys. Spectral signatures showed instances of substantial spectral overlap between habitats, causing classification confusion. Spectral separability between eelgrass and unvegetated cover diminished at the transition from shallow to deep water, creating an erroneous eelgrass boundary between unvegetated habitat and deep water in both of our North Harbour models. However, this was not detected in the confusion matrix, which showed eelgrass classification errors stemming from unvegetated seabed and macroalgae. Therefore, we suggest that model accuracy estimates may be misleading and enforce the importance of refining/validating RPAS surveys with complimentary ground-level surveys. Furthermore, consistent with previous studies (e.g.,
Barrell and Grant 2015;
Duffy et al. 2018;
Nahirnick et al. 2019b), sparse habitats or edges were more difficult to differentiate than continuous habitat. This may be because habitats underwater are more likely to appear different in aerial imagery due to light refraction (
Ellis et al. 2020;
Joyce et al. 2018). Additionally, classification errors are more likely to occur at habitat boundaries, where spectral signatures of boundary pixels are composed of a mixture of habitat types (
Woodcock and Strahler 1987), which generates pixels with intermediate tones, confusing the classification model. Thus, our results suggest that reproducible RPAS surveys of either deep (>5 m), fragmented, or more heterospecific eelgrass meadows are more difficult to acquire and seasonal estimates of change may be more prone to error. Separating submerged aquatic vegetation into eelgrass and macroalgae classes likely explains the lower model classification accuracies in our study. Using multispectral sensors may facilitate the detection of submerged benthic habitats in estuarine environments (
James et al. 2020;
Su et al. 2006), but this is beyond the scope of our study.
Various post-processing workflows have been applied in the literature to classify remote sensing imagery (e.g.,
Murfitt et al. 2017;
Duffy et al. 2018;
Ventura et al. 2018;
Wilson et al. 2019;
Schroeder et al. 2019). Most approaches include some form of manual classification to delineate features of interest or to create training data
post-hoc based on the analyst’s interpretation of the system and imagery (e.g.,
Konar and Iken 2018;
Ventura et al. 2018;
Gonçalves et al. 2019;
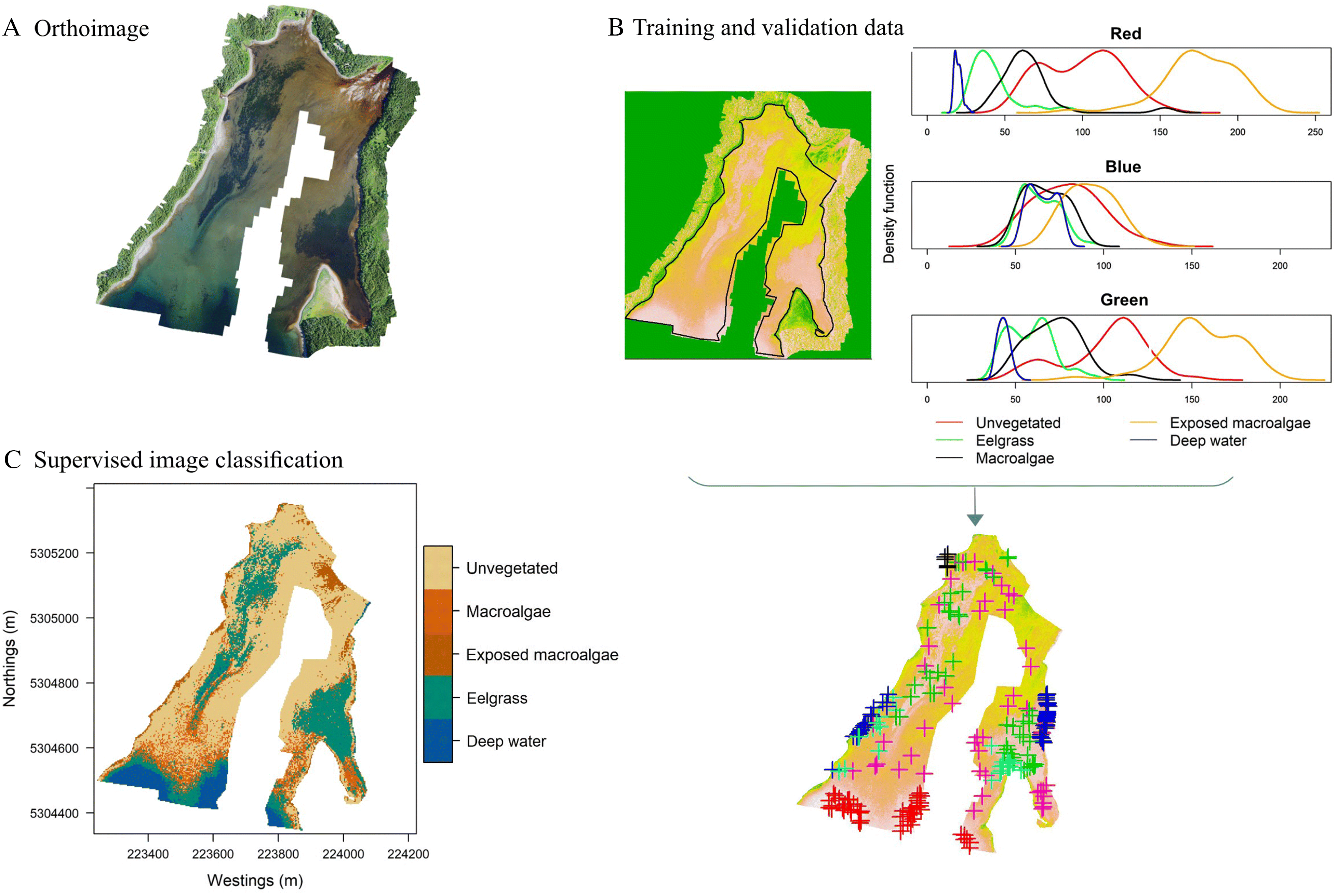
Nahirnick et al. 2019b). This introduces observer bias into the classification process, impacting the reproducibility of the survey. We present a method using kernel density plots to supplement training and validation data
post-hoc, which minimizes analyst-induced bias. However, some subjectivity was still required to select training data, such as ensuring these data do not come from pixels falling on habitat boundaries (
Lillesand and Kiefer 2000), and avoiding clustering training data to avoid overfitting sections of the model. Our study only tested the reproducibility of RPAS surveys using RGB imagery without supplemental information. Supplementing training data with additional habitat-defining characteristics such as depth and optical texture or collecting multispectral imagery to separate habitats (e.g.,
Tait et al. 2019) would likely improve the reproducibility of RPAS surveys for monitoring submerged eelgrass meadows and adjacent estuarine habitat and may be required when reproducibility is low.
Do results improve with altitude?
Terrestrial and coastal RPAS surveys have shown that lower altitude surveys enable more habitats to be distinguished in the imagery (
Duffy et al. 2018;
Perroy et al. 2017). However, we found that reducing survey altitude did not always improve agreement in estimates submerged eelgrass and macroalgae percent cover between RPAS surveys and snorkel quadrat surveys. Full site surveys showed that the accuracy of RPAS estimates improved with altitude for the edge transects, while it was unaffected for the inside transects. This could be because finer details such as ripples and shadows on the surface were detected by lower altitude images, increasing spectral variability (
Mount 2005;
Woodcock and Strahler 1987) and potentially reducing accuracy. For continuous eelgrass meadows, such disturbances are likely minimal on inside transects at lower altitudes given that the pixels are from the same habitat type. In contrast, the edge of the eelgrass meadow is more fragmented. Therefore, intermediate spectral tones from multiple habitats in a pixel and water disturbance will more likely distort habitat boundaries, altering apparent eelgrass coverage. At a higher altitude, noise from texture features such as ripples would have been reduced as lower resolution images smooth finer details and reduce spectral variability (
Woodcock and Strahler 1987). Therefore, the accuracy of RPAS surveys of meadow edges or of fragmented meadows may be improved in higher altitude surveys.
In contrast, the quadrat altitude test suggested that the effect of increasing altitude varied among habitats. Similar results were reported by
Ellis et al. (2020) while flying a fixed wing RPAS at similar altitudes as our study. Reduced image resolution with higher altitude increases the number of pixels that encompass a mixture of habitat types, and thus the number of pixels with intermediate color tones, which subsequently reduces classification accuracy (
Woodcock and Strahler 1987). We could therefore expect that, in heterogenous meadows, higher altitude would decrease classification accuracy. While this was the case for eelgrass and macroalgae, classification of unvegetated habitat was less affected by changes in image resolution with changing altitude. This is likely because eelgrass and macroalgae have similar spectral signatures regardless of resolution, while unvegetated habitat was more spectrally separable from the other two habitat classes. Nevertheless, in all cases the effect of increasing altitude only changed the estimates of cover by up to 10%. Flying at higher altitudes helps survey larger scale meadows within limited battery life of the RPAS and reduces the risk of environmental conditions that change within minutes (e.g., sun angle, tide, cloud cover) from changing during the survey (
Joyce et al. 2018). Therefore, in the case of large, submerged eelgrass meadows, the benefit of surveying at a higher altitude may outweigh the cost of collecting lower resolution imagery, especially given that images collected at lower altitudes can be more challenging to stitch into an orthomosaic.
Conclusions
Seasonal monitoring of seagrass meadow extent and adjacent estuarine habitats is important for coastal habitat management. In Newfoundland, seasonal changes in seagrass extent and cover would be more likely to be detected with survey periods starting earlier in the spring (e.g., April/May) after ice-melt and when eelgrass cover is low, then in the late summer/early fall (e.g., August/September). Issues of ice formation in the late fall/winter when eelgrass die-off (i.e., December) and increased riverine discharge due to snow melt in the spring, in addition to other varying environmental conditions (e.g., tide, wind, cloud), render seasonal monitoring using RPAS extremely challenging in north temperate environments. For these reasons, monitoring using RPAS in this region is optimal in summer/early fall months (i.e., July-early October), when eelgrass is the least fragmented (i.e., peak growth) and environmental conditions are more conducive to RPAS surveys. Similar logic could be applied to other temperate regions as well.
Regardless of the monitoring period, recognizing the limitations of RPAS surveys is crucial for reliable monitoring. We show that sources of variability on RPAS surveys limit reproducibility and therefore limit how much RPAS can be relied on to detect fine-scale changes during management decision-making, including changes in percent cover of seagrass and adjacent benthic habitats. In our study, trends in seasonal changes in seagrass and macroalgae percent cover often differed between RPAS and snorkel transect surveys. Differences in tidal height may have contributed to the variability observed; however, this is a significant challenge and potentially a limitation if RPAS are to be used to monitor submerged coastal habitats in temperate regions, where low wind and no cloud conditions are rare. Supplementing imagery with texture data such as bathymetry maps, Secchi depth, or multispectral imagery could help correct for depth and potentially help overcome reproducibility challenges but come at an additional cost and may be unfeasible for many monitoring groups. Overall, selecting whether to use RPAS surveys to monitor seagrass over time is question dependent. RPAS appear to be reliable for large-scale surveys to detect macro-habitat characteristics, such as general extent and identifying meadow structure and delineating vegetated versus unvegetated habitat in general. For example, edges and fragmented areas should be monitored via snorkel while large, submerged seagrass meadows can be monitored using higher altitude RPAS imagery. In this case, increasing survey altitude helps mitigate the risk of environmental conditions shifting mid-survey, thereby offsetting the cost of obtaining lower resolution imagery. Meanwhile, RPAS equipped with RGB sensors alone cannot distinguish between macroalgae and seagrass reliably, and estimates of fine-scale habitat changes, such as changes in species composition or eelgrass growth through time, should be interpreted with caution.