Structuring forces of intertidal infaunal communities on the northern coast of British Columbia, Canada: assessing the relative importance of top-down, bottom-up, middle-out, and abiotic variables
Abstract
While trophic and habitat-related abiotic variables (predation, competition, tolerance, etc.) are known to influence community structure in many ecosystems, some systems appear to be only minimally influenced by these variables. Sampling multiple tidal flat communities in northern BC, Canada, we investigated the relative importance of top-down and middle-out (mesopredators) variables, competition for resources (bottom up), and abiotic variables in structuring an infaunal community (invertebrates living in sediment). Similar to previous studies on mudflats in the Bay of Fundy (also at a north temperate latitude), we determined that these variables accounted for a minor (0%–9%) proportion of the observed variation in this infaunal community, suggesting that these variables play a small role in structuring this community. Based on the results of our study and in combination with previous experiments on infaunal recovery patterns post disturbance, we posit that the main factors influencing these infaunal communities likely operate at a scale of sites (kilometres) and(or) plot (metres or less) but not transects (10–100 m within site). Candidate forces structuring these intertidal communities that need future examination include regional species pools and the variables that affect these pools, sediment biogeochemistry, and disturbance/recovery history of a site. The similarity of our Pacific coast findings to those from the north temperate Atlantic coast suggests some similarity in the processes structuring these distinct infaunal communities.
Introduction
An important goal in ecology is the development of general theories that can explain observed spatiotemporal variation in biological communities (Pearson and Rosenberg 1978; Hubbell 2005; Huston 2014). These then enhance our ability to predict outcomes of natural or anthropogenic disturbances (Pearson and Rosenberg 1978; Orwin et al. 2016), as well as of restoration following disturbance (Campbell et al. 2019a). Frameworks for theory on forces underlying community composition and dynamics include niche vs. neutral dynamics (Hubbell 2005) and the designation of such forces into categories related to habitat availability, species availability, and species performance once in the habitat (such as predation, competition, and tolerance to environmental conditions) (Pickett et al. 1987; Meiners et al. 2015), among other frameworks.
Here, we examine possible species performance forces (predation, competition, and response to certain abiotic conditions) underlying community structure. For instance, environmental or abiotic variables, such as water salinity, temperature, or sediment properties, coupled with variations in exhibited tolerances of organisms for these conditions (Stillman 2002; Lu et al. 2008; Dashtgard et al. 2014; Sizmur et al. 2019) can play an important role in structuring communities (Kelaher et al. 2001; Ferguson et al. 2013; Gerwing et al. 2022a). As well, interactions among the living components of an ecosystem can affect communities, namely top-down predation, which in our analysis, and as discussed below, may be indistinguishable from bioturbation (Heck Jr. and Valentine 2007; Hughes et al. 2014; Johnson et al. 2014) and bottom-up forces, such as the availability and competition for nutrients (Davis et al. 2014; Schuldt et al. 2014; Pilditch et al. 2015a). Many communities appear shaped by a combination of top-down and bottom-up forces (Hamilton et al. 2006; Bracken et al. 2014; Greenville et al. 2014). Added complexity is observed in systems with mesopredators (mid-trophic level predators) (Prugh et al. 2009), which act both as predator and prey (Ambrose Jr. 1991; Marczak et al. 2011). These species can exert strong structuring influences upon a community via predation, bioturbation, and competition in a middle-out manner (Elmhagen and Rushton 2007; Quijón and Snelgrove 2008; Cunningham et al. 2020). Designing studies to elucidate the individual role of abovementioned forces in structuring a community is relatively straightforward; however, it is more difficult to quantify the relative importance of these forces, occurring concurrently, in determining community structure and dynamics (Wootton 1994; Agrawal et al. 2007; Dray et al. 2012; Gerwing et al. 2022b).
While trophic and habitat-related abiotic variables are known to influence community structure in many ecosystems, some systems appear to be only minimally influenced by them. For instance, ephemeral freshwater habitats, such as floodplains, can act as resource-pulse ecosystems (Crook et al. 2020; Nelson et al. 2021). In such systems, amply available resources, whose availability is temporally limited, trigger a cascading response throughout the ecosystem, minimizing the controlling influence of competition for resources, predation, and an individual species’ preference for abiotic conditions (Tilman 2004; Letnic and Dickman 2010; Greenville et al. 2014). In resource-pulse systems, more research is required to better understand how these communities are structured. Another study, pertinent to our current study, focused on intertidal mudflats in the upper Bay of Fundy (NB and NS, Canada). Gerwing et al. (2016) assessed the relative importance of top-down, bottom-up, middle-out, and sediment-related abiotic variables in structuring the infaunal community (invertebrates living in the sediment). They observed that while top-down, bottom-up, middle-out, and abiotic variables accounted for a statistically significant proportion of the observed community variation (1%–11%), the majority of that variation (79%) was accounted for by spatial factors, such as study site or patch.
Gerwing et al. (2016) suggested two sets of hypotheses for their observed patterns. First, high abundance of resources on the expansive Bay of Fundy mudflats may both minimize bottom-up control (e.g., by limiting competition for nutrients from sediment organic matter and benthic diatoms) and support a sufficiently dense population of potential prey items to dilute the importance of top-down and middle-out predation/bioturbation. Furthermore, the relatively low importance of abiotic variables in structuring the infaunal community could be a result of generally low horizontal physical heterogeneity observed on their study sites, as well as high nutrient concentrations that limit the influence of abiotic factors by attracting animals to patches with abiotic characteristics that would normally reduce occupancy. Pulses of amply available resources may result in intertidal mudflats operating as a resource-pulse ecosystem. Secondly, Gerwing et al. (2016) also hypothesized that since mudflats are a relatively benign environment for organisms adapted to living in mud, the inhabiting communities were not predominantly structured by top-down, bottom-up, middle-out, or abiotic factors. Instead, these communities may reflect priority effects or a “first come, first served” process (Sutherland 1974; Connell and Slatyer 1977). Initial delivery of larvae or colonization by juveniles and adults may play a predominant role in explaining the observed spatiotemporal variation in this type of community (Snelgrove et al. 1999; Pilditch et al. 2015a;2015b). In contrast to the resource-pulse hypothesis, priority effects imply some competition for resources.
While these sets of hypotheses are intriguing, there are questions about their applicability to other tidal flat systems. The Bay of Fundy is home to the largest tides in the world, a phenomenon that results in considerable amounts of sediment being suspended in the water column each day (Wu et al. 2011). Sediment dynamics may be partly responsible for the relatively limited biodiversity exhibited on the upper Bay of Fundy mudflats, by filtering out species unable to survive in such situations. For instance, the community of these Bay of Fundy mudflats is composed of roughly 10 infaunal taxa, 3 top-down predators, and 3 mesopredators (Gerwing et al. 2015a). Conversely, mudflat communities on the Pacific coast of BC, Canada, are composed of roughly 39 infaunal taxa, 8 top-down consumers, and 12 mesopredators (Campbell et al. 2020). Given the unique characteristics of Bay of Fundy mudflats, it is unclear whether the hypotheses of Gerwing et al. (2016) can be applied to other mudflat or tidal flat communities. Moreover, field studies focused on infaunal community recovery from anthropogenic and natural disturbances along the Pacific and Atlantic coasts suggest that priority effects may not be the predominant force structuring these communities (Norris et al. 2022). Instead, community trajectories may be strongly influenced by species availability, particularly regional species pools (Beukema et al. 1999; Thrush et al. 2003; Gerwing et al. 2017a; Campbell et al. 2019a; Cox et al. 2019; Gerwing et al. 2022c; Norris et al. 2022). As such, more work is required to better understand the suite of forces structuring infaunal communities.
Our goal for the present paper was to use community and population level correlations to investigate the potential relative importance of top-down, bottom-up, middle-out, and abiotic variables in structuring the infaunal community of tidal flats (both sand and mudflats) of the north coast of BC (Supplementary Table S1). We hypothesized that, as observed in Gerwing et al. (2016), top-down, bottom-up, middle-out, and abiotic variables would not associate well with the infaunal community. If top-down, bottom-up, middle-out, and abiotic variables have only a minor influence in structuring tidal flat communities on the Pacific coast, as inferred by Gerwing et al. (2016) on the Atlantic coast, it is possible that similar processes are operating in other infaunal communities elsewhere, thus expanding our empirical understanding of the relative importance of the forces that structure soft-sediment intertidal communities.
Methods
Study sites
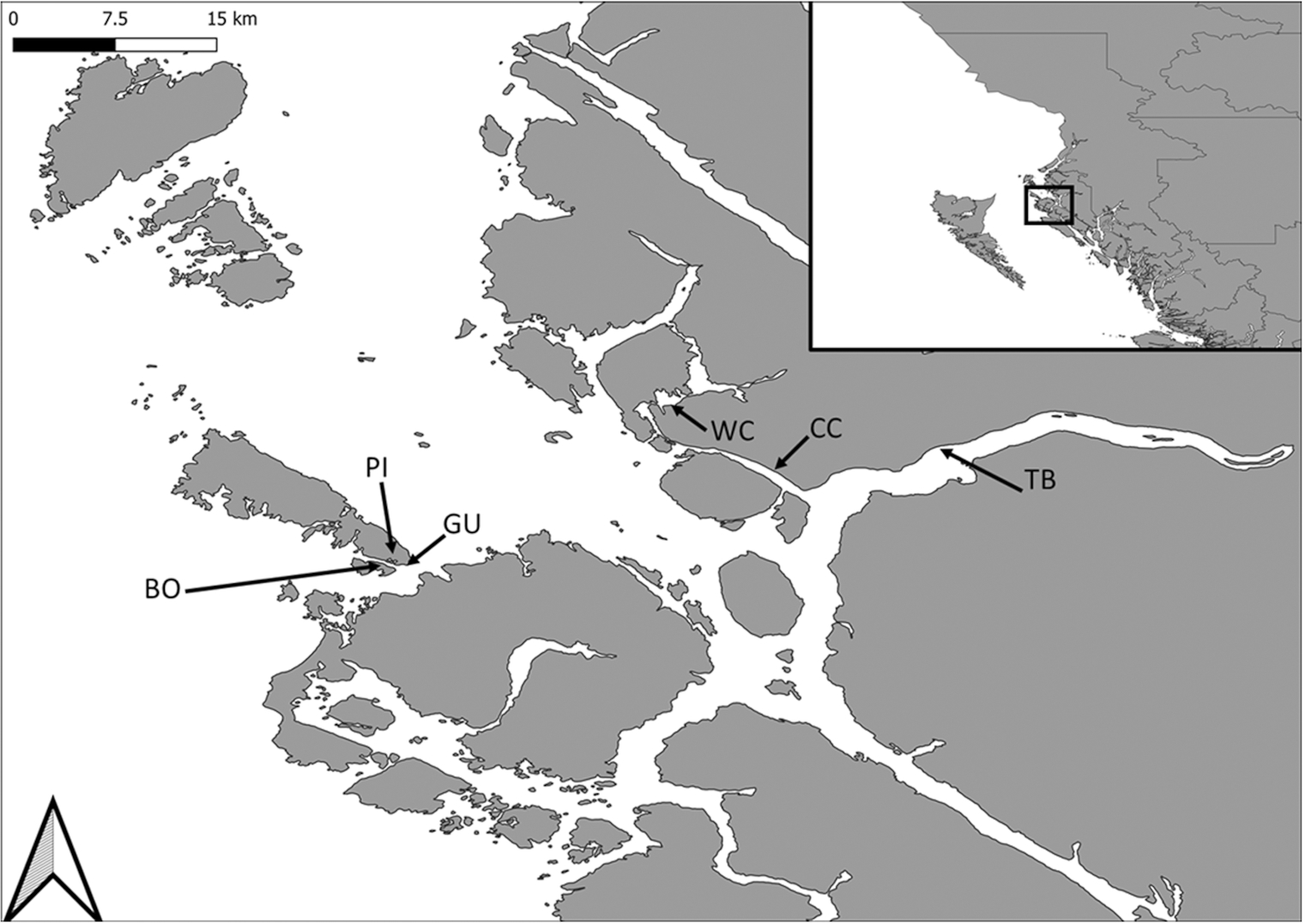
Six tidal flats (termed sites) were examined along the north coast of BC (Fig. 1), near the Skeena River (WC, Wolfe Cove; CC, Cassiar Cannery; TB, Tyee Bank; BO, Boulder Beach; PI, Prescott Inlet; GU, Coast Guard Beach). Typical amplitude of the semi-diurnal tides is 5–6.5 m. Sediment at WC, CC, and TB is primarily composed of silt and clay (mud), while that at BO, PI, and GU has a higher predominance of sand (Campbell et al. 2020; Gerwing et al. 2020a); amounts of gravel and cobble are low. Additional details about the sites are provided in Supplementary Table S1. The intertidal area in the region has a history of disturbance associated with human development, particularly from pulp mill effluent (operations ceased in 2001). Despite this history, tidal flats have naturally recovered from these disturbances (though indications of previous disturbance remain), with top layers of sediment (∼20 cm) currently uncontaminated by metal pollutants and host to an abundant and diverse invertebrate community (Campbell et al. 2019a, 2019b; Sizmur et al. 2019; Gerwing et al. 2020b). Our tidal flats are not main foraging areas for migrating shorebirds, such as the Western Sandpipers (Calidris mauri (Cabanis, 1857)); however, small flocks of 10–100 birds are observed in spring (T.G. Gerwing, personal observation).
Fig. 1.
Sampling design
At each site, 5 randomly selected transects (60–200 m long; depending on mudflat across-shore width) were established running from the landward start of the tidal flat to the low water line, separated by ∼20–100 m. Transects were stratified into equisized zones based on distance from shore (near, middle, and far). Within each zone, one sampling location was randomly selected (n =3 per transect, 15 per site). Sites were sampled four times in summer 2017 (Round A: 23 May–1 June, Round B: 21–26 June, Round C: 19–25 July, Round D: 18–24 August; 15 samples per site per round) on the lowest low tides of the year for a total of 60 samples per site and 360 total over all rounds. More details of the sampling design can be found in Campbell et al. (2020).
Biotic variables
At each sampling location, a 1 m2 plot was established to count epifauna (invertebrates living on the sediment), within which infauna were collected using a sediment core (10 cm length and 7 cm in diameter). For the latter, the sediment was passed through a 250 µm sieve, and the content stored in vials with 95% ethanol for later identification (Campbell et al. 2020). A pit (20 cm long, 20 cm wide, and 20 cm deep) was also dug in the plot where the core was taken to sample and identify larger or more mobile specimens that may have been missed by the infaunal core; these individuals were scarce and were integrated with abundances from core samples in the estimation of densities (standardized to numbers per m2). All infaunal specimens were identified to the lowest possible taxonomic unit as follows: cumaceans, amphipods, tanaids, polychaetes, nemerteans, and bivalves to species; chironomid larvae to family; copepods to order; ostracods to class; and nematodes to phylum (Campbell et al. 2020). Variable taxonomic resolution does not impair contrasts between sites, as Gerwing et al. (2020b) showed that analyzing infauna community data with specimens identified to different taxonomic levels produced similar results. In addition, a small sediment core (2–3 mm deep, 2 cm diameter) was taken adjacent to the infaunal core to measure abundance of benthic diatoms by determining concentration of chlorophyll a. Chlorophyll a pigment was extracted from sediment samples using buffered acetone, and its reflectance quantified using a spectrophotometer (Coulthard and Hamilton 2011).
Abiotic variables
At each sampling location, sediment properties were also measured. Sediment penetrability, an integrative variable that reflects the overall in situ physical conditions experienced by biota (Meadows et al. 1998; Gerwing et al. 2020a), was assessed by dropping a metal weight (15 cm long, 1.9 cm diameter, 330 g) from a height of 0.75 m above the sediment and measuring how far it penetrated into the sediment. Higher penetrability indicates finer-grained sediment with high water content, with few rocks or shell hash present in or on the sediment. Lower penetrability is indicative of larger-grained sediment with low water content, with more rocks or shell hash present (Hsu et al. 2009; Gerwing et al. 2020a). Water content (drying at 110 °C for 12 h), organic matter content (ashing at 550 °C for 4 h), and volume-weighted average particle size in the sediment’s top 1 cm were measured from sediment cores (5 cm depth, 4.5 cm diameter), collected adjacent to the abovementioned infaunal cores; more details of these processes can be found in Campbell et al. (2020). In the field, the void created in the sediment from the corer to collect infauna was used to measure depth to the apparent redox potential discontinuity, aRPD (Gerwing et al. 2013). aRPD depth is a relative measure of sediment porewater dissolved oxygen and redox conditions. Sediment with a deeper aRPD has more available dissolved oxygen than that with a shallower aRPD depth (Gerwing et al. 2018b). The proportion of each 1 m2 plot covered in woody debris, as well as deposited algae and eelgrass (Zostera spp.) debris, was also quantified, as this debris can create hypoxic conditions and smother infauna. Salinity was measured at the seaward end of each transect on each tidal flat on each sampling trip with a YSI multimeter in the water approximately 1 cm above the sediment surface at high tide. Finally, relative distance of each plot from the start of the mudflat (transition of saltmarsh, bedrock, or sandy beaches to mud (Gerwing et al. 2016)) was used as a proxy for intertidal elevation or duration of inundation for each plot location.
Data analysis
Analyses were conducted in the program PRIMER V7 (Clarke and Gorley 2015), with the PERMANOVA add on (Anderson et al. 2008). A permutational multivariate analysis of covariance (ANCOVA) (PERMANCOVA; 9999 permutations) was used to quantify the relationship between the infaunal community and top-down, bottom-up, middle-out, and abiotic variables (Gerwing et al. 2016). The multivariate response was a resemblance matrix of the densities of 39 taxa of infaunal invertebrates, calculated using Bray-Curtis similarity. Taxa densities were fourth root transformed to better consider the influence of abundant and rare species on the outcome of the analysis. A dummy variable of 0.01, below our threshold of detection, was added to address plots with no observed infauna (Anderson et al. 2008). Within the PERMANCOVA, round (four levels), site (six levels), and transect nested within site (five transects per site) were included as random factors and hereafter referred to as structural variables (since they reflect the structure of our sampling design).
Top-down, bottom-up, middle-out, and abiotic variables were included as covariates. Top-down predators (or bioturbators) included eight species whose diet could include infaunal taxa, or who would influence infauna through bioturbation (Pagliosa 2005; Light 2007; Jumars et al. 2015), but were unlikely to experience high levels of predation themselves during low tide (snails, crabs, and hermit crabs: Littorina sitkana Philippi, 1846; Littorina scutulata Gould, 1849; Cancer productus Randall, 1840; Cancer magister Dana, 1852; Hemigrapsus oregonensis (Dana, 1851); Pagurus hirsutiusculus (Dana, 1851); Pagurus granosimanus (Stimpson, 1859); and Hemigrapsus nudus (Dana, 1851). Middle-out variables, or mesopredators, were composed of four species (ribbon worms and sea cucumbers: Emplectonema gracile (Johnston, 1837); Paranemertes peregrina Coe, 1901; Leptosynapta albicans (Selenka, 1867); and Tubulanus polymorpha Renier, 1804, which are predominantly found on the sediment surface and may act as both predator and prey (Light 2007). This category does not include eight species of infaunal polychaete mesopredators (Hemigrapsus oregonensis; Eteone californica Hartman, 1936; Alitta brandti Malmgren, 1865; Scoletoma zonata (Johnson, 1901); Nephtys caeca (Fabricius, 1780); Nephtys caecoides Hartman, 1938; Glycera macrobranchia Moore, 1911; and Exogone lourei Berkeley & Berkeley, 1938), which in this analysis were classified as part of the infaunal community to better understand this community as a whole. We did this because these infaunal mesopredators could not be included both as covariates and in the multivariate response variable. However, these predatory polychaete worms were included as covariates in the subsequent individual species analyses detailed below. We also included two nutrient-based resources as bottom-up variables (chlorophyll a concentration and sediment organic matter content) and seven abiotic variables [sediment penetrability, water content, volume-weighted mean particle size, aRPD depth, water salinity, and possible causes of hypoxic conditions (% cover by algae and eelgrass)].
Prior to analysis, we assessed possible correlations between all pairs of covariates by calculating univariate Pearson’s correlation coefficients. We used a threshold of 0.95 (Clarke and Ainsworth 1993) for variables too correlated to be considered independent. Since the highest correlation coefficient observed was 0.76, all variables were included in our models. All covariates (except relative distance from shore) were fourth root transformed to correct for skewed distributions and then normalized.
As part of the PERMANCOVA, we quantified components of variation, the proportion of the multivariate variation accounted for by each independent variable (Searle et al. 1992; Anderson et al. 2008; Gerwing et al. 2016). An α of 0.05 was used to determine statistical significance for all analyses (Beninger et al. 2012). Finally, covariates that did not account for any variation in the multivariate response were removed (Fletcher and Underwood 2002; Gerwing et al. 2016).
Since multivariate analyses can obscure key taxa-specific relationships (Spasojevic and Suding 2012; Murray et al. 2014; Gerwing and Hawkes 2021), a PERMANCOVA (which also works well as a univariate analysis) was conducted for selected infaunal taxa. This selection was conducted via similarity percentages analyses (SIMPER; Clarke 1993), which identify taxa contributing the most to observed community variation between sites and rounds. Only taxa that accounted for ≥5% of the observed community variation between sites in the SIMPER analysis were retained for subsequent univariate analyses (Americorophium salmonis (Stimpson, 1857); Capitella species complex; Cumella vulgaris Hart, 1930; Eteone californica; Exogone lourei; Harpacticoida, Leptochelia spp.; Macoma balthica (Linnaeus, 1758); Nematoda, Nippoleucon hinumensis (Gamo, 1967); Nutricola tantilla (Gould, 1853); Oligochaeta, Ostracoda, Pygospio elegans Claparède, 1863; and S. zonata). Resemblance matrices were constructed and PERMANCOVA performed as detailed above with one exception. In taxa-specific PERMANOCOVA, all middle-out variables could be included (11–12 species). Univariate Pearson’s correlation test was then calculated to determine the nature of the relationship between covariates and a given response taxon (+/−).
Results
Structural factors accounted for most of the observed variation in the infaunal community (81%), with site (our largest spatial scale, 35%) and plot (our smallest spatial scale, 33%) accounting for the most (Table 1). Transect, which was intermediate in spatial scale and integrated across intertidal depth, accounted only for ∼3% of the community variation. While most of the relationships central to our study were statistically significant, top-down consumers only accounted for 0.4% of the variation, middle-out predators 0.2%, and abiotic variables 9% (Table 1). The covariate that accounted for the highest proportion of the variation was water salinity (7%). Bottom-up variables (nutrients) did not have a significant influence.
Table 1.
| Category | Source | df | MS | Pseudo-F | Unique permutations | p | Variance components (%) |
|---|---|---|---|---|---|---|---|
| Top down (0.40%) | Cancer magister | 1 | 1361.10 | 1.34 | 9957 | 0.25 | 0.06 |
| Littorina sitkana | 1 | 1617.00 | 1.09 | 9944 | 0.39 | 0.02 | |
| Hemigrapsus nudus | 1 | 3574.50 | 2.66 | 9944 | 0.02 | 0.40 | |
| Mesopredators (0.19%) | Emplectonema gracile | 1 | 2203.30 | 1.89 | 9959 | 0.08 | 0.19 |
| Paranemertes peregrina | 1 | 1636.90 | 1.56 | 9956 | 0.17 | 0.10 | |
| Leptosynapta albicans | 1 | 5596.00 | 1.60 | 9948 | 0.15 | 0.45 | |
| Pagurus granosimanus | 1 | 1984.40 | 1.77 | 9946 | 0.14 | 0.15 | |
| Nutrients (0%) | Organic matter content | 1 | 21 447.00 | 1.74 | 9950 | 0.12 | 2.59 |
| Chlorophyll a | 1 | 5414.00 | 1.17 | 9957 | 0.35 | 0.15 | |
| Abiotic (9.17%) | Penetrability | 1 | 8290.30 | 2.25 | 9952 | 0.04 | 1.18 |
| aRPD depth | 1 | 2912.80 | 2.17 | 9950 | 0.04 | 0.29 | |
| Water content | 1 | 1759.60 | 1.31 | 9949 | 0.27 | 0.49 | |
| Sediment particle size | 1 | 18 025.00 | 1.53 | 9951 | 0.18 | 2.72 | |
| Water salinity | 1 | 79 861.00 | 2.25 | 9941 | 0.03 | 7.70 | |
| Algal cover | 1 | 32 780.00 | 1.82 | 9939 | 0.11 | 2.83 | |
| Eelgrass cover | 1 | 3760.50 | 1.36 | 9946 | 0.23 | 0.18 | |
| Structural (80.51%) | Site | 5 | 19 578.00 | 9.83 | 9857 | 0.0001 | 35.34 |
| Round | 3 | 5343.30 | 3.62 | 9929 | 0.0001 | 3.49 | |
| Transect (site) | 24 | 1132.80 | 2.31 | 9787 | 0.0001 | 3.53 | |
| Site × round | 14 | 1581.20 | 3.26 | 9842 | 0.0001 | 4.86 | |
| Round × transect (site) | 72 | 485.06 | 0.90 | 9665 | 0.89 | 0.00 | |
| Residual (a.k.a. plot) | 225 | 539.16 | 33.29 | ||||
| Total | 359 |
Note: Significant sources of variation are in bold (α = 0.05). The % in parentheses is the sum of the statistically significant components of variation associated with that category.
Individual taxa exhibited a similar trend, with the majority of the variation accounted for by structural factors (Table 2). However, several key exceptions were noted, where covariates accounted for ≥10% of the observed variation in an infaunal taxon. These exceptions were correlations between water salinity and Cumella vulgaris (16%), Leptochelia spp. (28%), Oligochaeta (11%), Nematoda (12%), and Nutricola tantilla (15%); sediment organic matter content correlated with Nippoleucon hinumensis (22%) and Capitella species complex (12%); algal cover correlated with S. zonata (31%) and Exogone lourei (17%); and sediment particle size correlated with Macoma balthica (17%). Finally, a mixture of positive and negative correlations was observed between covariates and individual populations (Table 2).
Table 2.
| Category | Variables | Americorophium salmonis | Cumella vulgaris | Nippoleucon hinumensis | Leptochelia spp. | Eteone californica | Scoletoma zonata | Exogone lourei | Pygospio elegans | Capitella species complex | Oligochaete | Nematoda | Harpacticoida | Ostracoda | Macoma balthica | Nutricola tantilla |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Top Down | Cancer productus | 0.59% (+) | ||||||||||||||
| Hemigrapsus oregonensis | 0.23% (−) | |||||||||||||||
| Pagurus hirsutiusculus | 0.91% (+) | |||||||||||||||
| Pagurus granosimanus | 0.64% (−) | 1.1% (−) | ||||||||||||||
| Hemigrapsus nudus | 0.37% (+) | |||||||||||||||
| Mesopredator | Glycinde picta | 9.64% (+) | ||||||||||||||
| Scoletoma zonata | 2.68% (+) | 6.45% (−) | 4.87% (−) | |||||||||||||
| Eteone californica | 1.12% (−) | |||||||||||||||
| Nephtys caeca | 2.07% (+) | 4.39% (+) | ||||||||||||||
| Nephtys caecoides | ||||||||||||||||
| Glcyera macrobranchia | 2.05% (−) | 0.54% (−) | ||||||||||||||
| Exogone lourei | 2.08% (−) | 22.59% (−) | ||||||||||||||
| Paranemertes peregrina | 1.70% (+) | |||||||||||||||
| Leptosynapta albicans | 1.31% (+) | |||||||||||||||
| Nutrients | Organic matter content | 6.85% (+) | 22.03% (+) | 12.00% (+) | ||||||||||||
| Chlorophyll a | 1.39% (−) | |||||||||||||||
| Abiotic | Penetrability | 1.55% (−) | ||||||||||||||
| aRPD | 0.99% (+) | 1.00% (+) | 1.44% (−) | |||||||||||||
| Salinity | 16.05% (+) | 28.23% (+) | 10.68% (+) | 12.21% (+) | 14.53% (+) | |||||||||||
| Algae cover | 30.88% (+) | 17.38% (+) | ||||||||||||||
| Eelgrass cover | 0.79% (+) | |||||||||||||||
| Sediment particle size | 4.15% (−) | 16.92% (−) | ||||||||||||||
| Structural | Round | 3.19% | 25.57% | |||||||||||||
| Site | 9.63% | 20.81% | 34.45% | 45.36% | 15.37% | 25.44% | 21.62% | 18.78% | 16.35% | 15.57% | 13.12% | 8.29% | 18.04% | |||
| Transect (site) | 6.12% | 0.65% | 2.18% | 6.15% | 5.63% | 3.37% | ||||||||||
| Round × site | 16.72% | 10.12% | 21.45% | 0.91% | 3.94% | 9.24% | 5.46% | |||||||||
| Round × transect (site) | 7.40% | |||||||||||||||
| Residual (a.k.a plot) | 61.68% | 39.81% | 39.13% | 20.13% | 41.87% | 20.09% | 34.63% | 52.94% | 62.27% | 50.60% | 58.65% | 64.53% | 77.40% | 40.94% | 57.36% |
Discussion
For tidal flat systems on the north coast of BC, we attempted to elucidate the relative importance of top-down, bottom-up, middle-out, and abiotic variables in potentially structuring an infaunal community. In general, we observed that these variables accounted for a minority of the observed variation in this community; however, some differences were observed between individual taxa and community-level results.
Dominance of structural factors on observed community variation
Overall, spatial structural factors, namely site and plot, accounted for the majority (81%) of our observed variation in the infaunal community (Table 1; Fig. 2). As in other studies, the infaunal community varied significantly with top-down (Heck Jr. and Valentine 2007; Hughes et al. 2014), middle-out (Prugh et al. 2009; Johnson et al. 2014), and abiotic variables (Lu et al. 2008; Dashtgard et al. 2014), but these variables accounted for a minority of the community variation (0%–9% each; 10% total). The overall low proportion of the variation accounted for by these covariates does not mean that these variables are unimportant in structuring the infaunal community, since our analyses use correlational data to report on relative importance and not on absolute effect size. Other studies have observed that top-down, bottom-up, middle-out, and environmental variables may exert structuring influences upon infaunal communities at limited spatiotemporal scales (Commito and Ambrose Jr. 1985; Wilson 1990; Lindsay et al. 1996; Williams et al. 2004; Hamilton et al. 2006; Cheverie et al. 2014; Gerwing et al. 2022b). However, our findings suggest that at broad spatiotemporal scales, infaunal communities are relatively uncoupled from the controlling influences of top-down, bottom-up, middle-out, and a number of sediment abiotic variables.
Fig. 2.

In habitats where predation and attendant bioturbation are common, if the prey base is large enough or predation/bioturbation is patchy, predation and bioturbation can have a minor effect on the prey community (Tilman 2004; Letnic and Dickman 2010; Greenville et al. 2014). Community dynamics that are relatively independent from predation/bioturbation (top down or middle out) have been observed before in infaunal communities (Gerwing et al. 2016), as well as in resource-pulse ecosystems. In the latter ecosystems, large increases in prey populations stimulated by pulses of resources result in a prey community so large that predators exert little influence over the density or dynamics of the prey species (Gray et al. 2002; Greenville et al. 2014; Crook et al. 2020). Tidal flats (at least in north temperate latitudes) experience regular pulses in nutrients in the form of benthic diatoms and organic matter, mainly during spring and summer (de Jong and de Jonge 1995; Gerwing et al. 2015a; Campbell et al. 2020; Schnurr et al. 2020). These resource pulses result in a rapid increase in infaunal density and richness. We posit that these increases are so large that predation and bioturbation, both top down and middle out (epifaunal and mesopredators), have a minor impact upon this infaunal community (Gerwing et al. 2016). While some predators/bioturbators may have localized and short-term effects upon certain taxa (Drolet et al. 2009; Cheverie et al. 2014), induced mortality is likely compensatory (Pöysä 2004). Abundances in infaunal communities located in north temperate latitudes, like ours, decline naturally each year as winter approaches (Reise 1991; Gerwing et al. 2015a; Campbell et al. 2020). Therefore, predators are likely consuming soon-to-die individuals (compensatory mortality, sensu (Hamilton et al. 2006)), further minimizing the impact of predation upon these populations and communities.
Infaunal community structure and dynamics were also uncoupled from availability of nutrients (chlorophyll a concentration and sediment organic matter content; Table 1). High primary productivity of intertidal flats in spring and summer (Hargrave et al. 1983; Gerwing et al. 2015a; Campbell et al. 2020) likely limits the role exploitative competition plays in structuring the infaunal community, a relationship that is also commonly observed in resource-pulse ecosystems (Letnic and Dickman 2010; Greenville et al. 2014; Crook et al. 2020).
Abiotic variables mainly related to sediment properties and salinity only accounted for 9% of the infaunal community variation (Table 1). Gerwing et al. (2016) observed a similar phenomenon and partially attributed this observation to the limited range of sediment abiotic conditions observed in their study sites. However, our study included a broader range and variation of abiotic conditions (both between and within sites; Supplementary Table S1). Moreover, the most important abiotic covariate in our study was water salinity, which accounted for 8% of the infaunal community variation. The influence of salinity upon invertebrate community structure is well known (Rosenberg et al. 1992; Owen and Forbes 1997; Rutger and Wing 2006; Lu et al. 2008). Despite this variable not being included within Gerwing et al.’s (2016) analysis, our study reported a similar influence of abiotic variables upon the infaunal community (Table 3; 9%–11%). Thus, it is unlikely that the low proportion of the community variation accounted for by abiotic variables (related to tidal flat environments) is a product of low variability in sediment conditions or missing variables within Gerwing et al.’s (2016) analysis.
Table 3.
| Variables | Pacific coast | Atlantic coast |
|---|---|---|
| Top down | 0.4% | 6% |
| Middle out | 0.5%* | 1%* |
| Bottom up | 0% | 1% |
| Abiotic | 9% | 11% |
| Structural | 81% | 79% |
*
Middle-out variables are averaged (from Table 2 in the present study and Table 2 in Gerwing et al. (2016)), since mesopredators are also a component of the infaunal community, and so had to be analyzed separately.
Structural factors, which accounted for 81% of the observed variation in the infaunal community (Table 1), are challenging to quantify. For instance, the temporal term round (3%) may represent interactions between the time of year and weather or climate patterns (Scholz and Liebezeit 2012; Drolet et al. 2013b). In our study, plots (1 m2) accounted for 33% of the observed community variation. This factor may represent localized hydrology and delivery of larvae, post-settlement dispersal, small-scale (within-plot) unmeasured intra- and interspecific interactions, and(or) availability of biogeochemical elements in the sediment (Drolet et al. 2013a; Pilditch et al. 2015b; Sizmur et al. 2019; Gerwing et al. 2022a, 2022b). Site, representing spatial variation on the scale of kilometres, is likely a product of processes such as larval supply (Weersing and Toonen 2009), post-settlement dispersal (Pilditch et al. 2015b), as well as unmeasured site variables such as hydrology, biogeochemistry, exposure to waves and tidal currents (Williams et al. 2013; Gerwing et al. 2015b; Rubin et al. 2017; Gerwing et al. 2022a), or site-specific histories, such as previous disturbance and human development (Gerwing et al. 2017c; Campbell et al. 2019a; Sizmur et al. 2019).
Patterns at the level of an individual taxon
In general, a similar pattern was observed when this infaunal community was analyzed as a whole and as individual taxa: the majority of the variation was accounted for by spatial structural factors (Tables 1 and 2; Fig. 2). This further supports the idea that these infaunal species are mostly uncoupled from the controlling influence of top-down, bottom-up, middle-out, and abiotic variables. In particular, none of the measured top-down variables accounted for more than 1% of the observed variation of an individual infaunal taxon (Table 2). When combined with the results from the community-level analysis, it appears that top-down variables may not play a major role in structuring these infaunal taxa, nor the community as a whole.
Nevertheless, some stronger relationships were noted, and these identify areas for further research, as well as potentially useful indicator species. For example, Capitella species complex, a sessile polychaete worm, was positively associated with sediment organic matter content (12%; Table 2). This relationship was not unexpected since this capitellid family is a known indicator of organic enrichment, often found thriving in areas with elevated levels of sediment organic matter (Pearson and Rosenberg 1976, 1978; Gerwing et al. 2018a; Campbell et al. 2019a). As such, this observation suggests that these polychaetes can be used as indicators of organic enrichment in this Pacific intertidal system as well.
With regard to abiotic variables, water salinity had a positive correlation with five taxa: Cumella vulgaris (16%), Leptochelia spp. (28%), Oligochaeta (11%), Nematoda (12%), and Nutricola tantilla (15%) (Table 2). As mentioned above, the influence of salinity upon infaunal community composition and density is well reported (Rosenberg et al. 1992; Owen and Forbes 1997; Rutger and Wing 2006; Lu et al. 2008). In our study, the positive correlations between these taxa and salinity suggest that these taxa increase in density the further away the intertidal site was from the influence of the Skeena River’s freshwater input. Finally, a negative correlation was observed between Macoma balthica (17%) and sediment particle size, indicating that in our study area, M. balthica was more common on flats composed of mud than sand (Campbell et al. 2020). This observation has been made before (Cranford et al. 1985; Gerwing et al. 2015a, 2015b).
While middle-out variables may be relatively unimportant in structuring our infaunal community as a whole, at the level of individual taxa, we detected certain interactions that may be important to individual species at fine spatiotemporal scales. We observed 13 correlations between mesopredators and individual infaunal taxa, accounting for 1%–23% of infaunal population variation (Table 2). This suggests that mesopredators may play a minor but meaningful role in structuring infaunal communities and populations, a conclusion that has been reached by other studies of intertidal infauna (Ambrose Jr. 1984a, 1984b; Commito and Ambrose Jr. 1985; Commito and Shrader 1985; Ambrose Jr. 1991; Gerwing et al. 2022b). However, only one correlation was observed to exceed 10% of the infaunal variation: a negative correlation between S. zonata and Exogone lourei (23%). Interestingly, this relationship was unidirectional. When we assessed the variables associated with the spatiotemporal variation of S. zonata, a strong negative correlation with Exogone lourei was observed. However, when we assessed the variables associated with the spatiotemporal variation of Exogone lourei, no correlation with S. zonata was observed. This relationship could be the result of predation or competition, and more research is required to better elucidate the nature of this interaction.
Other hypotheses on predominant structuring forces of infaunal communities in tidal flats
Early studies examining forces that influence community variation in soft-sediment marine ecosystems (including tidal flats) questioned the importance of local interactions (i.e., species performance drivers) (Grassle and Sanders 1973; Thistle 1981). Based upon their recent work in the Bay of Fundy mudflats, Gerwing et al. (2016) also reasoned that intertidal infaunal communities are not predominantly structured by top-down, bottom-up, middle-out, or sediment variables (considered species performance forces). Despite being conducted in tidal flats that exhibited greater biological diversity and abiotic differences, the results of our current study mirrored those of Gerwing et al. (2016). Such similarity in results from disparate study sites on the Atlantic and Pacific coasts of Canada suggests some similarity in the main structuring forces of these infaunal communities. It would be interesting to re-examine whether other tidal flat infaunal communities in north temperate latitudes, such as those in the Wadden Sea (Flach and Beukema 1995) and the UK (Waldock et al. 1999), are structured in a similar manner. Overall, these results continue to raise questions about the primary drivers of infaunal community structure.
Our data suggest that unmeasured or unidentified spatial terms, operating at the site or plot level, may be the dominant structuring factors. Gerwing et al. (2016) postulated that intertidal infaunal communities may be structured by a first-come-first-served process (priority effects) following a disturbance, as can be observed in rocky shores when recruitment is low (Sutherland 1974; Bertness 2007; Fukami 2015). Under such a process, delivery of larvae and dispersing propagules as well as movement by juveniles and adults (Stocks 2002; Drolet et al. 2012; Bringloe et al. 2013; Pilditch et al. 2015b) may play important structuring roles at the spatial scale of the plot (metres) and site (hundreds of metres to kilometres) following the disturbance. This process could explain the large proportion of the variation accounted for by structural terms in our results (Tables 1 and 2), as well as in the results of Gerwing et al. (2016) (Table 3). Once individuals are established, they may be able to resist colonization by subsequent dispersing of individuals via pre-emptive competition or inhibition (Ambrose Jr 1984a; Loeuille and Leibold 2008), resulting in priority effects dominating infaunal community structure and dynamics.
However, studies of infaunal community recovery from anthropogenic and natural disturbances along the Pacific and Atlantic coasts of Canada suggest that priority effects may not be the predominant force structuring these communities (Norris et al. 2022). Instead, community trajectories appeared to be strongly influenced by species availability, particularly regional species pools, and not by the order of species arrival. Following disturbance and then establishment, infaunal community structure (species composition and abundances) increased in similarity to the surrounding areas (Beukema et al. 1999; Thrush et al. 2003; Gerwing et al. 2017a; Campbell et al. 2019a; Cox et al. 2019; Gerwing et al. 2022c; Norris et al. 2022). Regional species pools have been implicated in structuring other marine invertebrate communities (Weslawski 2003; Witman et al. 2004; Somerfield et al. 2009; de Juan and Hewitt 2011), and we posit that they may play a substantial role in structuring intertidal infaunal communities both on small and large scales.
Another set of hypotheses about drivers of infaunal community structure, related to species performance, that we only superficially mention in our study is the modification of the sedimentary environment by infauna (Reise 2002; Woodin et al. 2010). While our study mostly considered trophic-related variables, infauna can also stabilize sediments (e.g., bioconstruction), destabilize sediments (including bioturbation), cause bioadvection of porewater (affecting local distribution of dissolved oxygen and various other chemicals) while undertaking all sorts of behaviours, and subsequently facilitate or inhibit other community members (Rhoads and Young 1970; Reise 2002; Woodin et al. 2010; Gerwing et al. 2017b). Sediment geochemistry (whether mediated by biota or not) could affect communities at the patch level or site level, where the presence/absence and concentrations of key elements within the sediment may influence infaunal survival, physiology, behaviour, and abundance (Volkenborn et al. 2012; Sizmur et al. 2013; Chiarelli and Roccheri 2014; Sizmur et al. 2019; Gerwing et al. 2022a). More research is required to evaluate these various hypotheses related to non-trophic species performance (such as facilitation), species availability (such as the regional species pool), and habitat availability (such as disturbance type, scale, and frequency) on tidal flats in northern BC as well as in the Bay of Fundy.
Conclusions
Using intertidal soft-sediment communities in northern BC, we investigated relationships among top-down, bottom-up, middle-out, and abiotic variables with infaunal community composition. As in Gerwing et al.’s (2016) work in a similar ecosystem in the Bay of Fundy, we observed that these forces accounted for a minor proportion of the observed variation in infaunal community, suggesting a minor role in structuring this community. We posit that regular pulses of nutrient-related resources on tidal flats enable them to act like resource-pulse ecosystems. Where freely available and abundant, these resources contribute to limiting the effects of top-down, bottom-up, middle-out, and abiotic variables as structuring forces. Gerwing et al. (2016) also hypothesized that priority effects (pre-emptive competition) may be a primary force structuring infaunal communities. However, we now suggest, based on our present study in combination with recent studies on recovery of the infaunal community following various disturbances (Thrush et al. 2003; Gerwing et al. 2017a, 2018a; Cox et al. 2019; Sizmur et al. 2019; Campbell et al. 2019a; Gerwing et al. 2022c; Norris et al. 2022) that colonizers and subsequent infaunal community composition may reflect regional species pools and the variables that influence these pools. More research is required to evaluate this hypothesis.
Acknowledgements
Field work was done with assistance from D Parker, J Starr, and Z Starr. Funding was provided by the Kitsumkalum First Nation to TGG, a Natural Sciences and Engineering Research Council (NSERC) of Canada Engage grant to TGG and FJ, a MITACS Elevate grant, partnered with the Hakai Institute, to TGG, and an NSERC CGS M to LC. Cassiar Cannery provided further support to TGG and LC. We dedicate this paper to the memory of Mark Bell. Wherever you are Mark, we know that the beer is cold, the fishing is good, and the boats are made of cedar. Sleep well my friend.
References
Agrawal A.A., Ackerly D.D., Adler F., Arnold A.E., Cáceres C., Doak D.F., et al. 2007. Filling key gaps in population and community ecology. Frontiers in Ecology and the Environment, 5: 145–152.
Ambrose W.G. Jr. 1984a. Influence of residents on the development of a marine soft-bottom community. Journal of Marine Research, 42: 633–654.
Ambrose W.G. Jr. 1984b. Influences of predatory polychaetes and epibenthic predators on the structure of a soft-bottom community in a Maine estuary. Journal of Experimental Marine Biology and Ecology, 81: 115–145.
Ambrose W.G. Jr. 1991. Are infaunal predators important in structuring marine soft-bottom communities? American Zoologist, 31: 849–860.
Anderson M., Gorley R.N., Clarke R.K. 2008. Permanova+ for primer: guide to software and statistical methods. PRIMER-E Ltd, Plymouth, UK.
Beninger P.G., Boldina I., Katsanevakis S. 2012. Strengthening statistical usage in marine ecology. Journal of Experimental Marine Biology and Ecology, 426: 97–108.
Bertness M.D. 2007. Atlantic shorelines: natural history and ecology. Princeton University Press, Princeton, NJ.
Beukema J., Flach E., Dekker R., Starink M. 1999. A long-term study of the recovery of the macrozoobenthos on large defaunated plots on a tidal flat in the wadden sea. Journal of Sea Research, 42: 235–254.
Bracken M.E.S., Dolecal R.E., Long J.D. 2014. Community context mediates the top-down versus bottom-up effects of grazers on rocky shores. Ecology, 95: 1458–1463.
Bringloe T.T., Drolet D., Barbeau M.A., Forbes M.R., Gerwing T.G. 2013. Spatial variation in population structure and its relation to movement and the potential for dispersal in a model intertidal invertebrate. PLoS ONE, 8: e69091.
Campbell L., Sizmur T., Juanes F., Gerwing T.G. 2019a. Passive reclamation of soft-sediment ecosystems on the north coast of British Columbia, canada. Journal of Sea Research, 155: 101796.
Campbell L., Wood L., Allen Gerwing A.M., Allen S., Sizmur T., Rogers M., et al. 2019b. A rapid, non-invasive population assessment technique for marine burrowing macrofauna inhabiting soft sediments. Estuarine, Coastal and Shelf Science, 227: 106343.
Campbell L., Dudas S.E., Juanes F., Allen Gerwing A.M., Gerwing T.G. 2020. Invertebrate communities, sediment parameters, and food availability of intertidal soft-sediment ecosystems on the north coast of British Columbia, Canada. Journal of Natural History, 54: 919–945.
Cheverie A.V., Hamilton D.J., Coffin M.R.S., Barbeau M.A. 2014. Effects of shorebird predation and snail abundance on an intertidal mudflat community. Journal of Sea Research, 92: 102–114.
Chiarelli R., Roccheri M.C. 2014. Marine invertebrates as bioindicators of heavy metal pollution. Open Journal of Metal, 4: 93–106.
Clarke K.R. 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology, 18: 117–143.
Clarke K.R., Ainsworth M. 1993. A method of linking multivariate community structure to environmental variables. Marine Ecology Progress Series, 92: 205–219.
Clarke K.R., Gorley R.N. 2015. Primer v7: user manual/tutorial. Primer-E Ltd, Plymouth, UK.
Commito J.A., Ambrose W.G. Jr. 1985. Multiple trophic levels in soft-bottom communities. Marine Ecology Progress Series, 26: 289–293.
Commito J.A., Shrader P. 1985. Benthic community response to experimental additions of the polychaete Nereis virens. Marine Biology, 86: 101–107.
Connell J.H., Slatyer R.O. 1977. Mechanisms of succession in natural communities and their role in community stability and organization. The American Naturalist, 111: 1119–1144.
Coulthard M.E., Hamilton D.J. 2011. Effects of Ilyanassa obsoleta (say) on the abundance and vertical distribution of Corophium volutator (pallas) on mudflats of the upper Bay of Fundy. Journal of Experimental Marine Biology and Ecology, 397: 161–172.
Cox K.D., Gerwing T.G., Macdonald T., Hessing-Lewis M., Millard-Martin B., Command R.J., et al. 2019. Infaunal community responses to ancient clam gardens. ICES Journal of Marine Science, 76: 2362–2373.
Cranford P., Peer D., Gordon D. 1985. Population dynamics and production of Macoma balthica in Cumberland Basin and Shepody Bay, Bay of Fundy. Netherlands Journal of Sea Research, 19: 135–146.
Crook D.A., Buckle D.J., Morrongiello J.R., Allsop Q.A., Baldwin W., Saunders T.M., Douglas M.M. 2020. Tracking the resource pulse: movement responses of fish to dynamic floodplain habitat in a tropical river. Journal of Animal Ecology, 89: 795–807.
Cunningham C.X., Johnson C.N., Jones M.E. 2020. A native apex predator limits an invasive mesopredator and protects native prey: Tasmanian devils protecting bandicoots from cats. Ecology Letters, 23: 711–721.
Dashtgard S.E., Pearson N.J., Gingras M.K. 2014. Sedimentology, ichnology, ecology and anthropogenic modification of muddy tidal flats in a cold-temperate environment: Chignecto Bay, Canada. Geological Society, 388: 229–245.
Davis B., Mattone C., Sheaves M. 2014. Bottom-up control regulates patterns of fish connectivity and assemblage structure in coastal wetlands. Marine Ecology Progress Series, 500: 175–186.
De Jong D., De Jonge V.N. 1995. Dynamics and distribution of microphytobenthic chlorophyll-a in the Western Scheldt estuary (SW Netherlands). Hydrobiologia, 311: 21–30.
De Juan S., Hewitt J. 2011. Relative importance of local biotic and environmental factors versus regional factors in driving macrobenthic species richness in intertidal areas. Marine Ecology Progress Series, 423: 117–129.
Dray S., Pélissier R., Couteron P., Fortin M.-J., Legendre P., Peres-Neto P.R., et al. 2012. Community ecology in the age of multivariate multiscale spatial analysis. Ecological Monographs, 82: 257–275.
Drolet D., Barbeau M.A., Coffin M.R.S., Hamilton D.J. 2009. Effect of the snail Ilyanassa obsoleta (say) on dynamics of the amphipod Corophium volutator (pallas) on an intertidal mudflat. Journal of Experimental Marine Biology and Ecology, 368: 189–195.
Drolet D., Bringloe T.T., Coffin M.R.S., Barbeau M.A., Hamilton D.J. 2012. Potential for between-mudflat movement and metapopulation dynamics in an intertidal burrowing amphipod. Marine Ecology Progress Series, 449: 197–209.
Drolet D., Coffin M.R.S., Barbeau M.A., Hamilton D.J. 2013a. Influence of intra-and interspecific interactions on short-term movement of the amphipod Corophium volutator in varying environmental conditions. Estuaries and Coasts, 36: 1–11.
Drolet D., Kennedy K., Barbeau M.A. 2013b. Winter population dynamics and survival of the intertidal mudflat amphipod Corophium volutator (pallas). Journal of Experimental Marine Biology and Ecology, 441: 126–137.
Elmhagen B., Rushton S.P. 2007. Trophic control of mesopredators in terrestrial ecosystems: top-down or bottom-up? Ecology Letters, 10: 197–206.
Ferguson N., White C.R., Marshall D.J. 2013. Competition in benthic marine invertebrates: the unrecognized role of exploitative competition for oxygen. Ecology, 94: 126–135.
Flach E.C., Beukema J.J. 1995. Density-governing mechanisms in populations of the lugworm Arenicola marina on tidal flats. Oceanographic Literature Review, 42.
Fletcher D.J., Underwood A.J. 2002. How to cope with negative estimates of components of variance in ecological field studies. Journal of Experimental Marine Biology and Ecology, 273: 89–95.
Fukami T. 2015. Historical contingency in community assembly: integrating niches, species pools, and priority effects. Annual Review of Ecology, Evolution, and Systematics, 46: 1–23.
Gerwing T.G., Hawkes V.C. 2021. Similarity analyses in restoration ecology and how to improve their utility. Restoration Ecology, 29: e13368.
Gerwing T.G., Allen Gerwing A.M., Drolet D., Hamilton D.J., Barbeau M.A. 2013. Comparison of two methods of measuring the depth of the redox potential discontinuity in intertidal mudflat sediments. Marine Ecology Progress Series, 487: 7–13.
Gerwing T.G., Allen Gerwing A.M., Drolet D., Barbeau M.A., Hamilton D.J. 2015a. Spatiotemporal variation in biotic and abiotic features of eight intertidal mudflats in the upper Bay of Fundy, Canada. Northeastern Naturalist, 22: 1–44.
Gerwing T.G., Drolet D., Barbeau M.A., Hamilton D.J., Allen Gerwing A.M. 2015b. Resilience of an intertidal infaunal community to winter stressors. Journal of Sea Research, 97: 40–49.
Gerwing T.G., Drolet D., Hamilton D.J., Barbeau M.A. 2016. Relative importance of biotic and abiotic forces on the composition and dynamics of a soft-sediment intertidal community. PLoS ONE, 11: e0147098.
Gerwing T.G., Allen Gerwing A.M., Macdonald T., Cox K., Juanes F., Dudas S.E. 2017a. Intertidal soft-sediment community does not respond to disturbance as postulated by the intermediate disturbance hypothesis. Journal of Sea Research, 129: 22–28.
Gerwing T.G., Allen Gerwing A.M., Cox K., Juanes F., Dudas S.E. 2017b. Relationship between apparent redox potential discontinuity (aRPD) depth and environmental variables in soft-sediment habitats. International Journal of Sediment Research, 32: 472–480.
Gerwing T.G., Hamilton D.J., Barbeau M.A., Haralampides K.A., Yamazaki G. 2017c. Short-term response of a downstream marine system to the partial opening of a tidal-river causeway. Estuaries and Coasts, 40: 717–725.
Gerwing T.G., Campbell L., Allen Gerwing A.M., Allen S., Cox K., Rogers M., et al. 2018a. Potential impacts of logging on intertidal infaunal communities within the kitimat river estuary. Journal of Natural History, 52: 2833–2855.
Gerwing T.G., Cox K., Allen Gerwing A.M., Carr-Harris C.N., Dudas S.E., Juanes F. 2018b. Depth to the apparent redox potential discontinuity (aRPD) as a parameter of interest in marine benthic habitat quality models. International Journal of Sediment Research, 33: 472–480.
Gerwing T.G., Barbeau M.A., Hamilton D.J., Allen Gerwing A.M., Sinclair J., Campbell L., et al. 2020a. Assessment of sediment penetrability as an integrated in situ measure of intertidal soft-sediment conditions. Marine Ecology Progress Series, 648: 67–78.
Gerwing T.G., Cox K., Allen Gerwing A.M., Campbell L., Macdonald T., Dudas S.E., Juanes F. 2020b. Varying intertidal invertebrate taxonomic resolution does not influence ecological findings. Estuarine, Coastal and Shelf Science, 232: 106516.
Gerwing T.G., Allen Gerwing A.M., Davies M.M., Darcott K., Campbell L., Juanes F., et al. 2022a. Sediment geochemistry influences infaunal invertebrate community composition and population abundances. Marine Biology, 170.
Gerwing T.G., Campbell L., Thomson H.M., Dudas S.E., Juanes F. 2022b. Community and population impacts of an introduced meiofauna (Nippoleucon hinumensis) upon an intertidal infaunal community and its closest endemic analog (Cumella vulgaris). Aquatic Ecology, 57: 85–97.
Gerwing T.G., Dudas S.E., Juanes F. 2022c. Increasing misalignment of spatial resolution between investigative and disturbance scales alters observed responses of an infaunal community to varying disturbance severities. Estuarine, Coastal and Shelf Science, 265: 107718.
Grassle J.F., Sanders H.L. 1973. Life histories and the role of disturbance. Deep Sea Research and Oceanographic Abstracts, 20: 643–659.
Gray A., Simenstad C.A., Bottom D.L., Cornwell T.J. 2002. Contrasting functional performance of juvenile salmon habitat in recovering wetlands of the salmon river estuary, Oregon, USA. Restoration Ecology, 10: 514–526.
Greenville A.C., Wardle G.M., Tamayo B., Dickman C.R. 2014. Bottom-up and top-down processes interact to modify intraguild interactions in resource-pulse environments. Oecologia, 175: 1349–1358.
Hamilton D.J., Diamond A.W., Wells P.G. 2006. Shorebirds, snails, and the amphipod Corophium volutator in the upper Bay of Fundy: top-down vs. bottom-up factors, and the influence of compensatory interactions on mudflat ecology. Hydrobiologia, 567: 285–306.
Hargrave B.T., Prouse N.J., Phillips G.A., Neame P.A. 1983. Primary production and respiration in pelagic and benthic communities at two intertidal sites in the upper Bay of Fundy. Canadian Journal of Fisheries and Aquatic Sciences, 40: 229–243.
Heck K.L., Jr., Valentine J.F. 2007. The primacy of top-down effects in shallow benthic ecosystems. Estuaries and Coasts, 30: 371–381.
Hsu C.-B., Chen C.-P., Hsieh H.-L. 2009. Effects of sediment compaction on macroinfauna in a protected coastal wetland in Taiwan. Marine Ecology Progress Series, 375: 73–83.
Hubbell S.P. 2005. Neutral theory in community ecology and the hypothesis of functional equivalence. Functional ecology, 19: 166–172.
Hughes J.M., Stewart J.S., Lyle J.M., Suthers I.M. 2014. Top-down pressure on small pelagic fish by eastern Australian salmon Arripis trutta; estimation of daily ration and annual prey consumption using multiple techniques. Journal of Experimental Marine Biology and Ecology, 459: 190–198.
Huston M.A. 2014. Disturbance, productivity, and species diversity: empiricism vs. logic in ecological theory. Ecology, 95: 2382–2396.
Johnson K.D., Grabowski J.H., Smee D.L. 2014. Omnivory dampens trophic cascades in estuarine communities. Marine Ecology Progress Series, 507: 197–206.
Jumars P.A., Dorgan K.M., Lindsay S.M. 2015. Diet of worms emended: an update of polychaete feeding guilds. Annual review of marine science, 7: 497–520.
Kelaher B.P., Chapman M.G., Underwood A.J. 2001. Spatial patterns of diverse macrofaunal assemblages in coralline turf and their associations with environmental variables. Journal of the Marine Biological Association of the UK, 81: 917–930.
Letnic M., Dickman C.R. 2010. Resource pulses and mammalian dynamics: conceptual models for hummock grasslands and other Australian desert habitats. Biological Reviews, 85: 501–521.
Light S.F. 2007. The light and smith manual: intertidal invertebrates from central California to Oregon. University of California Press, Berkely, CA, USA.
Lindsay S.M., Wethey D.S., Woodin S.A. 1996. Modeling interactions of browsing predation, infaunal activity, and recruitment in marine soft-sediment habitats. The American Naturalist, 148: 684–699.
Loeuille N., Leibold M.A. 2008. Evolution in metacommunities: on the relative importance of species sorting and monopolization in structuring communities. The American Naturalist, 171: 788–799.
Lu L., Grant J., Barrell J. 2008. Macrofaunal spatial patterns in relationship to environmental variables in the Richibucto Estuary, New Brunswick, Canada. Estuaries and Coasts, 31: 994–1005.
Marczak L.B., Ho C.-K., Więski K., Vu H., Denno R., Pennings S. 2011. Latitudinal variation in top-down and bottom-up control of a salt marsh food web. Ecology, 92: 276–281.
Meadows P.S., Murray J.M., Meadows A., Wood D.M., West F.J. 1998. Microscale biogeotechnical differences in intertidal sedimentary ecosystems. Geological Society, 139: 349–366.
Meiners S.J., Cadotte M.W., Fridley J.D., Pickett S.T., Walker L.R. 2015. Is successional research nearing its climax? New approaches for understanding dynamic communities. Functional Ecology, 29: 154–164.
Murray F., Douglas A., Solan M. 2014. Species that share traits do not necessarily form distinct and universally applicable functional effect groups. Marine Ecology Progress Series, 516: 23–34.
Nelson K.R., Davies M.M., Thomson H.M., Lawn P., Kushneryk K., Brouard-John E., et al. 2021. Population dynamics and methodological assessments from a 15-year period of amphibian monitoring in British Columbia’s southern gulf islands. Environmental Monitoring and Assessment, 193: 1–12.
Norris G.S., Gerwing T.G., Barbeau M.A., Hamilton D.J. 2022. Using successional drivers to understand spatiotemporal dynamics in intertidal mudflat communities. Ecosphere, 13.
Orwin K.H., Wardle D.A., Towns D.R., John M.G.S., Bellingham P.J., Jones C., et al. 2016. Burrowing seabird effects on invertebrate communities in soil and litter are dominated by ecosystem engineering rather than nutrient addition. Oecologia, 180: 217–230.
Owen R., Forbes A. 1997. Salinity, floods and the infaunal macrobenthic community of the St Lucia Estuary, Kwazulu-Natal, South Africa. Southern African Journal of Aquatic Science, 23: 14–30.
Pagliosa P.R. 2005. Another diet of worms: the applicability of polychaete feeding guilds as a useful conceptual framework and biological variable. Marine Ecology, 26: 246–254.
Pearson T.H., Rosenberg R. 1976. A comparative study of the effects on the marine environment of wastes from cellulose industries in Scotland and Sweden. Ambio, 5: 77–79.
Pearson T.H., Rosenberg R. 1978. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanography and Marine Biology: An Annual Review, 16: 229–331.
Pickett S., Collins S., Armesto J.J. 1987. Models, mechanisms and pathways of succession. The Botanical Review, 53: 335–371.
Pilditch C.A., Leduc D., Nodder S.D., Probert P.K., Bowden D.A. 2015a. Spatial patterns and environmental drivers of benthic infaunal community structure and ecosystem function on the New Zealand continental margin. New Zealand Journal of Marine and Freshwater Research, 49: 224–246.
Pilditch C.A., Valanko S., Norkko J., Norkko A. 2015b. Post-settlement dispersal: the neglected link in maintenance of soft-sediment biodiversity. Biology letters, 11: 20140795.
Pöysä H. 2004. Ecological basis of sustainable harvesting: is the prevailing paradigm of compensatory mortality still valid? Oikos, 104: 612–615.
Prugh L.R., Stoner C.J., Epps C.W., Bean W.T., Ripple W.J., Laliberte A.S., Brashares J.S. 2009. The rise of the mesopredator. Bioscience, 59: 779–791.
QGIS. 2019. QGIS development team. QGIS geographic information system. Open source geospatial foundation project. Available from http://qgis.osgeo.org.
Quijón P.A., Snelgrove P.V. 2008. Trophic complexity in marine sediments: new evidence from the gulf of St. Lawrence. Marine Ecology Progress Series, 371: 85–89.
Reise K. (Editor). 1991. Mosaic cycles in the marine benthos. Springer.
Reise K. 2002. Sediment mediated species interactions in coastal waters. Journal of Sea Research, 48: 127–141.
Rhoads D.C., Young D.K. 1970. The influence of deposit-feeding organisms on sediment stability and community trophic structure. Journal of Marine Research, 28: 150–178.
Rosenberg R., Loo L.-O., Möller P. 1992. Hypoxia, salinity and temperature as structuring factors for marine benthic communities in a eutrophic area. Netherlands Journal of Sea Research, 30: 121–129.
Rubin S.P., Miller I.M., Foley M.M., Berry H.D., Duda J.J., Hudson B., et al. 2017. Increased sediment load during a large-scale dam removal changes nearshore subtidal communities. PloS ONE, 12: e0187742.
Rutger S.M., Wing S.R. 2006. Effects of freshwater input on shallow-water infaunal communities in doubtful sound, New Zealand.Marine Ecology Progress Series, 314: 35–47.
Schnurr P.J., Drever M.C., Elner R.W., Harper J., Arts M.T. 2020. Peak abundance of fatty acids from intertidal biofilm in relation to the breeding migration of shorebirds. Frontiers in Marine Science, 7: 63.
Scholz B., Liebezeit G. 2012. Microphytobenthic dynamics in a Wadden Sea intertidal flat–part ii: seasonal and spatial variability of non-diatom community components in relation to abiotic parameters. European Journal of Phycology, 47: 120–137.
Schuldt A., Baruffol M., Bruelheide H., Chen S., Chi X., Wall M., Assmann T. 2014. Woody plant phylogenetic diversity mediates bottom-up control of arthropod biomass in species-rich forests. Oecologia, 176: 171–182.
Searle S.R., Casella G., Mcculloch C.E. 1992. Variance components. John Wiley & Sons, New York.
Sizmur T., Canário J., Gerwing T.G., Mallory M.L., Oʼdriscoll N.J. 2013. Mercury and methylmercury bioaccumulation by polychaete worms is governed by both feeding ecology and mercury bioavailability in coastal mudflats. Environmental Pollution, 176: 18–25.
Sizmur T., Campbell L., Dracott K., Jones M., O’driscoll N.J., Gerwing T.G. 2019. Relationships between potentially toxic elements in intertidal sediments and their bioaccumulation by benthic invertebrates. PloS ONE, 14: e0216767.
Snelgrove P., Grant J., Pilditch C. 1999. Habitat selection and adult-larvae interactions in settling larvae of soft-shell clam Mya arenaria. Marine Ecology Progress Series, 182: 149–159.
Somerfield P., Arvanitidis C., Faulwetter S., Chatzigeorgiou G., Vasileiadou A., Amouroux J., et al. 2009. Assessing evidence for random assembly of marine benthic communities from regional species pools. Marine Ecology Progress Series, 382: 279–286.
Spasojevic M.J., Suding K.N. 2012. Inferring community assembly mechanisms from functional diversity patterns: the importance of multiple assembly processes. Journal of Ecology, 100: 652–661.
Stillman J.H. 2002. Causes and consequences of thermal tolerance limits in rocky intertidal porcelain crabs, genus Petrolisthes. Integrative and Comparative Biology, 42: 790–796.
Stocks K.I. 2002. Flume experiments on post-settlement movement in polychaetes. Journal of Marine Research, 60: 743–762.
Sutherland J.P. 1974. Multiple stable points in natural communities. American Naturalist, 108: 859–873.
Thistle D. 1981. Natural physical disturbances and communities of marine soft bottoms. Marine Ecology Progress Series, 6: 223–228.
Thrush S.F., Hewitt J.E., Norkko A., Cummings V.J., Funnell G.A. 2003. Macrobenthic recovery processes following catastrophic sedimentation on estuarine sandflats. Ecological Applications, 13: 1433–1455.
Tilman D. 2004. Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proceedings of the National Academy of Sciences of the United States of America, 101: 10854–10861.
Volkenborn N., Meile C., Polerecky L., Pilditch C.A., Norkko A., Norkko J., et al. 2012. Intermittent bioirrigation and oxygen dynamics in permeable sediments: an experimental and modeling study of three tellinid bivalves. Journal of Marine Research, 70: 794–823.
Waldock R., Rees H., Matthiessen P., Pendle M. 1999. Surveys of the benthic infauna of the crouch estuary (UK) in relation to TBT contamination. Journal of the Marine Biological Association of the United Kingdom, 79: 225–232.
Weersing K., Toonen R.J. 2009. Population genetics, larval dispersal, and connectivity in marine systems. Marine Ecology Progress Series, 393: 1–12.
Weslawski M.K.S.W.J. 2003. A multi-scale study of the biodiversity of the benthic infauna of the high-latitude Kongsfiord, Svalbard. Polar Biology, 26: 383–388.
Williams I., Van Der Meer J., Dekker R., Beukema J., Holmes S. 2004. Exploring interactions among intertidal macrozoobenthos of the Dutch Wadden Sea using population growth models. Journal of Sea Research, 52: 307–319.
Williams G.J., Smith J.E., Conklin E.J., Gove J.M., Sala E., Sandin S.A. 2013. Benthic communities at two remote pacific coral reefs: effects of reef habitat, depth, and wave energy gradients on spatial patterns. PeerJ, 1: e81.
Wilson W.H. 1990. Competition and predation in marine soft-sediment communities. Annual Review of Ecology and Systematics, 21: 221–241.
Witman J.D., Etter R.J., Smith F. 2004. The relationship between regional and local species diversity in marine benthic communities: a global perspective. Proceedings of the National Academy of Sciences, 101: 15664–15669.
Woodin S.A., Wethey D.S., Volkenborn N. 2010. Infaunal hydraulic ecosystem engineers: cast of characters and impacts.Integrative and Comparative Biology, 50: 176–187.
Wootton J.T. 1994. Predicting direct and indirect effects: an integrated approach using experiments and path analysis. Ecology, 75: 151–165.
Wu Y., Chaffey J., Greenberg D.A., Colbo K., Smith P.C. 2011. Tidally-induced sediment transport patterns in the upper Bay of Fundy: a numerical study. Continental Shelf Research, 31: 2041–2053.
Supplementary material
Supplementary Material 1 (DOCX / 62.9 KB).
- Download
- 62.94 KB
Information & Authors
Information
Published In

FACETS
Volume 8 • 2023
Pages: 1 - 14
Editor: Dr. S.J. Cooke
History
Received: 10 September 2022
Accepted: 6 February 2023
Version of record online: 4 June 2023
Copyright
© 2023 The Author(s). This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data is available upon reasonable request to the corresponding author.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: TGG, GSN, SED, FJ
Data curation: TGG, LC
Formal analysis: TGG
Funding acquisition: TGG, SED, FJ
Investigation: TGG, LC, DJH, MAB, GSN, SED, FJ
Methodology: TGG, LC
Project administration: TGG, LC, SED, FJ
Resources: TGG, LC, SED, FJ
Software: TGG, SED, FJ
Supervision: TGG, SED, FJ
Validation: TGG
Visualization: TGG
Writing – original draft: TGG
Writing – review & editing: TGG, LC, DJH, MAB, GSN, SED, FJ
Competing Interests
The authors have no competing interests or conflicts of interest to declare.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Travis G. Gerwing, Lily Campbell, Diana J. Hamilton, Myriam A. Barbeau, Gregory S. Norris, Sarah E. Dudas, and Francis Juanes. 2023. Structuring forces of intertidal infaunal communities on the northern coast of British Columbia, Canada: assessing the relative importance of top-down, bottom-up, middle-out, and abiotic variables. FACETS.
8: 1-14.
https://doi.org/10.1139/facets-2022-0199
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. Effects of Experimental Addition of Algae and Shell Hash on an Infaunal Mudflat Community