Age matters: Submersion period shapes community composition of lake biofilms under glyphosate stress
Abstract
The phosphonate herbicide glyphosate, which is the active ingredient in the commercial formulation Roundup®, is currently the most globally used herbicide. In aquatic ecosystems, periphytic biofilms, or periphyton, are at the base of food webs and are often the first communities to be in direct contact with runoff. Microcosm experiments were conducted to assess the effects of a pulse exposure of glyphosate on community composition and chlorophyll a concentrations of lake biofilms at different colonization stages (2 months, 1 year, and 20 years). This is the first study that uses such contrasting submersion periods. Biofilms were exposed to either environmental levels of pure analytical grade glyphosate (6 μg/L, 65 μg/L, and 600 μg/L) or to corresponding phosphorus concentrations. Community composition was determined by deep sequencing of the 18S and 16S rRNA genes to target eukaryotes and cyanobacteria, respectively. The results showed that submersion period was the only significant contributor to community structure. However, at the taxon level, the potentially toxic genus Anabaena was found to increase in relative abundance. We also observed that glyphosate releases phosphorus into the surrounding water, but not in a bioavailable form. The results of this study indicate that environmental concentrations of glyphosate do not seem to impact the community composition or metabolism of lake biofilms under pulse event conditions.
Graphical Abstract
Introduction
The phosphonate herbicide glyphosate, which is the active ingredient in Roundup®, is currently the most widely used herbicide in the world (Vera et al. 2010; Gomes et al. 2014; Vera et al. 2014; Gomes et al. 2016a, 2016b). In 2014, worldwide glyphosate use was 825 803 tons, 90.4% of which was used in an agricultural context (Benbrook 2016). The development of genetically modified crops designed to make them resistant to glyphosate has increased the use of the herbicide. Glyphosate is also used in urban and household settings, exacerbating its exposure potential (Benbrook 2016).
Glyphosate is a broad spectrum, non-selective systemic herbicide (Vera et al. 2010; Gomes et al. 2016b). Its principal mode of action is the inhibition of the synthesis of the 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), an enzyme involved in the biosynthesis of the essential aromatic amino acids phenylalanine, tyrosine, and tryptophan (Pérez et al. 2007; Gomes et al. 2014). Because of its short half-life, glyphosate can be rapidly degraded by microbes to aminomethylphosphonic acid (AMPA), a metabolite that can interfere with the biosynthesis of chlorophyll (Gomes et al. 2014). Glyphosate-based herbicides are typically sprayed on seedlings in the spring but may also be applied to crops prior to harvest (Benbrook 2016). This herbicide is, thus, subject to leaching into streams and rivers, leading to pulse exposure in aquatic organisms.
In aquatic ecosystems, periphytic biofilms, or periphyton, often contain photosynthetic organisms that contribute significantly to primary production in lakes and streams (Wetzel 1983; Wetzel 1993; Stevenson et al. 1996; Battin et al. 2016). They are also biogeochemical drivers of contaminant cycling, because they include phototrophic, heterotrophic, and even chemotrophic organisms (Desrosiers et al. 2006; Montuelle et al. 2010; Hamelin et al. 2011; Lázaro et al. 2013; Leclerc et al. 2015). These biofilms are a complex assemblage of living and dead bacteria, algae, and fungi embedded in a polysaccharide matrix that also includes organic and inorganic particles (Wetzel 1983). These communities are attached to submerged surfaces such as plants, rocks, and sediments. Periphytic biofilms are often the first communities to be in direct contact with runoff. Cyanobacteria are ubiquitous members of freshwater biofilms, and certain members of this bacterial phylum have the ability to degrade glyphosate and use it as a source of phosphorus, which constitutes 18.2% of glyphosate’s molecular weight (Lipok et al. 2007; Forlani et al. 2008; Vera et al. 2010). Biofilms are one of the most significant microbial communities at the base of food webs in shallow lakes (Vadeboncoeur and Steinman 2002). Thus, adverse effects on upper trophic levels could occur if the structure and function of these periphytic biofilms are negatively impacted by the toxic effects of glyphosate or by its use as a source of phosphorus. Glyphosate and AMPA can, therefore, alter freshwater food webs and fish stocks by affecting periphytic biofilms when they are leached into lakes and rivers.
At early colonization stages, biofilms can be seen as open systems with low levels of nutrients and carbon internal recycling. Early organisms, both autotrophs and heterotrophs, thus, remain dependent on the availability of nutrients in the water column (Hagerthey et al. 2011). At more mature colonization stages, microscopic ecological niches are created and nutrient exchanges with the water column decrease (Wetzel 1993; Hagerthey et al. 2011). In mature biofilms, communities are, therefore, more protected from external stressors (Tlili et al. 2011; Lozano et al. 2018). This protection can be explained by the fact that as colonization proceeds, the production of extracellular polymeric substances (EPS) by algae and bacteria, including polysaccharides, might increase, forming a barrier to the surrounding water (Ivorra et al. 2000). However, little is known about the importance of the colonization stages of biofilms in lakes. Studies have also shown that some pesticides and metals can alter the function and composition of periphytic communities, based on the assumption that, within biofilms, different species show distinct levels of sensitivity to these contaminants (Schmitt-Jansen and Altenburger 2005; Tlili et al. 2010; Lavoie et al. 2012). Even if some studies have assessed the effects of glyphosate on periphytic biofilms (Pérez et al. 2007; Vera et al. 2010; Vera et al. 2014; Lozano et al. 2018), its impact on community composition has mainly been evaluated through microscopic techniques (Pérez et al. 2007; Vera et al. 2010; Lozano et al. 2018; Smedbol et al. 2018).
The objective of this study was to identify the factors that shape the community composition of natural lake biofilms under a glyphosate pulse stress. We hypothesized that (1) mature biofilms would be less adversely impacted by glyphosate exposure because their exchanges with the water column are limited, and that (2) glyphosate exposure would shape community composition because of the varying sensitivities amongst species. To test these two hypotheses, microcosm experiments were conducted to assess the effects of glyphosate on the community composition of natural lake periphytic biofilms grown on artificial substrates at different stages of colonization (i.e., 2 months, 1 year, and 20 years). In addition to glyphosate treatments, some biofilms were also exposed to equivalent concentrations of phosphorus to compare the amount of phosphorus potentially available for algal growth in both types of exposure. Chlorophyll a content was used to assess the adverse impacts of glyphosate and community composition was determined using 18S and 16S rRNA gene analysis to target eukaryotes and cyanobacteria, respectively. To our knowledge, this was the first study to use such contrasting colonization stages, hereafter referred to as biofilm age.
Materials and methods
Study area and sampling
Biofilms were grown in situ on Teflon® artificial substrates in Lake Croche (0.179 km2; 45°59′N, 74°01′W), an oligotrophic Precambrian Shield lake (Desrosiers et al. 2006; Perron et al. 2014; Leclerc et al. 2015). This lake was considered to be free from prior pesticide exposure, as it is geographically isolated from agricultural lands. Thus, biofilms were not exposed to glyphosate during colonization. Substrate setups (Fig. S1) were installed as described by Desrosiers et al. (2006) and submerged for two months (19 July 2016–11 September 2016), 1 year (20 October 2015–21 September 2016) and 20 years (1996–2016), allowing differential colonization in terms of community structure and thickness. As Teflon is an inert substrate that favours exchanges between the biofilms and their environment, the use of such artificial substrates lessens perturbations of the community, allowing it to maintain its integrity (Desrosiers et al. 2006). Substrate setups were anchored with a clay brick at 1 m depth in the littoral zone. On the day of substrate harvesting, the colonized round mesh disks (9.6 cm2 surface area; 70 μm pore size) were removed from the supporting structure and kept in filtered (0.45 μm) lake water. Mesh disks and their biofilms were stored in the dark at low temperature during transport to the laboratory.
Experimental design
Three distinct pulse experiments were conducted for each different biofilm age (submersion periods of 2 months, 1 year, and 20 years). Pulse experiments were chosen over chronic exposure to mimic episodic runoff events following the application of glyphosate. For each experiment, colonized mesh disks were placed in round Pyrex® dishes (1-L capacity) filled with 600 mL of filtered (0.45 μm) lake water. Biofilms were acclimated to experimental conditions for 24 h before the addition of glyphosate or phosphorus. Biofilms were exposed to different initial concentrations of technical grade glyphosate PESTANAL® (Sigma-Aldrich, St. Louis, Missouri, USA); 99.7% purity; CAS: 1071-83-6) or phosphorus as KH2PO4 (Bregnard et al. 1996) for a period of 7 d (Fig. S2). Duration of exposure was chosen given preliminary results according to which only 30% of 2.5 mg/L glyphosate remained after 48 h under the same experimental conditions (Fig. S3). We chose to directly assess the effects of the active ingredient rather than a commercial formulation, because the latter contains a mixture of additives that can alter glyphosate’s toxicity (Vera et al. 2014). Treatments were performed in triplicate, for a total of 21 microcosms per experiment (Fig. S2). Experimental treatments are described in Table 1. Nominal initial concentrations of glyphosate were accurately measured in a preliminary study we conducted in 2015 under the exact same experimental conditions (Fig. S4). In a survey of Quebec streams and rivers, Giroux (2015) synchronized their sampling with the beginning of agricultural activities and the rainy season, as pesticides are subject to leaching to lakes and rivers following heavy rains. The highest glyphosate concentration they detected in an agricultural watershed was 6 μg/L. Because this concentration is much lower than most concentrations tested in the literature (Pérez et al. 2007; Vera et al. 2010; Vera et al. 2014; Smedbol et al. 2018), we chose it as our lowest glyphosate treatment level (Table 1).
Table 1.
| Treatment | Nominal concentration | Description |
|---|---|---|
| Control | Not applicable | No addition of glyphosate or phosphorus |
| G1 | 6 μg/L of glyphosate | Maximum concentration measured in the watershed of the Saint Lawrence River between 2011 and 2014 (Giroux 2015) |
| G2 | 65 μg/L of glyphosate | Water quality guideline for the protection of aquatic wildlife in Quebec (Giroux 2015) |
| G3 | 600 μg/L of glyphosate | 100 times the maximum environmental concentration |
| P1 | 1 μg/L of phosphorus | Concentration equivalent to the amount of phosphorus in G1 |
| P2 | 12 μg/L of phosphorus | Concentration equivalent to the amount of phosphorus in G2 |
| P3 | 110 μg/L of phosphorus | Concentration equivalent to the amount of phosphorus in G3 |
Note: Nominal initial concentrations of glyphosate were accurately measured in a preliminary study we conducted in 2015 under the exact same experimental conditions.
The microcosms were randomly distributed and incubated in a growth chamber at 23 °C, which was the temperature measured in the lake when biofilms were harvested on 31 August 2016 (average pH = 7.45 and conductivity = 9.5 μS/cm2). The day:night cycle was set to 14 h:10 h, according to illumination hours on this same day. These temperature and light parameters were maintained throughout the experiments. Constant oxygenation in the microcosms was ensured using an air pump system connected to air stones. We sampled water and periphyton from the microcosms at the very beginning of the experiment (T0), and after three (T3) and 7 d (T7).
Periphyton analysis
Two mesh disks were sampled from each microcosm on T0, T3, and T7. These two disks were brushed, pooled, and suspended in filtered (0.45 μm) lake water. Following stirring and homogenization, the suspension was subsampled and several parameters were measured.
Chlorophyll a (Chla)
For Chla analysis, 500 μL of the periphyton suspension was filtered on GF/F glass microfibre Whatman© filters (Whatman International Ltd., Maidstone, England) and kept frozen (−80 °C) until extraction with hot 90% ethanol (Nusch 1980). Extracts were kept at 4 °C overnight and absorbance values were measured before and after acidification (0.01 mL HCl 1 mol/L per 1 mL of extract) at 665 nm (A665) and 750 nm (A750) (Spectronic Unicam UV300 UV-Visible Spectrometer (Thermo Spectronic, Rochester, New York, USA)). Surface-normalized Chla concentrations (μg·cm−2) were calculated according to the following equation for each sample:where Ab = A665 – A750 before acidification, Aa = A665 – A750 after acidification, v = extraction volume (mL), E = total sample suspension volume/filtered subsample volume (L), L = spectrophotometer cell length (cm) and S = mesh disk surface that was brushed (cm2).
DNA extraction and sequence analyses
Nucleic acid extraction was performed on lyophilized biofilm samples using the DNeasy Plant Mini Kit (QIAGEN, Hilden, Germany) following the manufacturer’s instructions. Library preparation for 16S and 18S rRNA was done using Nextera XT (Illumina Inc., San Diego, California, USA), and next-generation sequencing was performed on an Illumina MiSeq (Illumina Inc., San Diego, California, USA). Reads were quality checked with FastQC (Andrews 2010), reads were filtered with FASTX-Toolkit (Gordon 2009) and BBMap (Bushnell 2014), and chimeras were removed with UCHIME (Edgar et al. 2011) in VSEARCH (Rognes et al. 2016). Operational taxonomic unit (OTU) picking (Greengenes version 13_8, 97% identity) was performed in QIIME (Caporaso et al. 2010). OTU tables were rarefied to 4000 and 10 000 sequences per sample for cyanobacteria-specific 16S and eukaryotic 18S, respectively. After quality filtering and rarefaction, we had 44 samples containing 916 OTUs for cyanobacteria and 49 samples containing 1920 OTUs for eukaryotes. Beta diversity analyses were performed with weighted and unweighted UniFrac (Lozupone and Knight 2005; Lozupone et al. 2011) as a distance metric in R (R Core Team 2016) using the phyloseq package (McMurdie and Holmes 2013). UniFrac uses distances between samples on a phylogenetic tree, either accounting for the relative abundance of OTUs (weighted) or not (unweighted). Community compositions among the different submersion periods were assessed. For full sequencing and read processing methods, see the Supplementary Methods (SM.1). Sequences were submitted into the Sequence Read Archive (SRA) database (National Center for Biotechnology Information (NCBI)) (SRA accession SRP150239, BioProject PRJNA475256).
Essential aromatic amino acids
Aromatic amino acids (phenylalanine, tryptophan, and serine) concentrations were measured in 20-year-old biofilms exposed to glyphosate for 7 d using an Agilent 6430 triple quadrupole (QQQ)-LC-MS/MS (Santa Clara, California, USA) in a preliminary experiment we performed in 2015. For a detailed description of the analysis, refer to the Supplementary Methods (SM.2).
Water analysis
Water was sampled in each microcosm for chemical analyses on T0 (immediately after glyphosate and phosphorus additions), T3, and T7. Glyphosate degradation was only determined in herbicide-treated microcosms.
Glyphosate
In 2015, we conducted the same experiment but with 20-year-old periphyton exclusively. During this preliminary study, analyses of glyphosate and AMPA were conducted by a provincial government laboratory. Glyphosate and AMPA were analyzed by liquid chromatography, post-column derivation, and fluorescence detection, with a detection limit of 0.08 μg/L for glyphosate and 0.4 μg/L for AMPA (CEAEQ method MA. 403—GlyAmp 1.0 2011-03-07) (CEAEQ 2008). In 2016, glyphosate enzyme-linked immunosorbent assay (ELISA) test kits were purchased from Abraxis LLC (Warminster, Pennsylvania, USA) and used following the manufacturer’s instructions. The method detection limit (MDL) and the limit of quantification (LOQ) as defined by Abraxis were 0.05 μg/L and 0.15 μg/L, respectively. Derivatized standard and control solutions were used every 12 samples to ensure analytical quality. Although the former analytical method allowed us to measure accurate glyphosate concentrations, we used the latter to show loss patterns.
Phosphorus
Total phosphorus (TP), dissolved phosphorus (DP), and orthophosphate levels were analyzed by flow injection analysis (Astoria2 Analyzer, Astoria-Pacific, Clackamas, Oregon, USA). Orthophosphates represent the phosphorus species that is most readily utilized by biota (Gaffney et al. 2001). Reactive phosphorus in our experiments was, thus, measured as orthophosphates, which provide a fair estimation of the amount of phosphorus available for algal growth (Strickland and Parsons 1972; Carignan and Neiff 1992). We assessed (1) the amount of reactive phosphorus that was released from glyphosate degradation, and (2) whether this reactive phosphorus decreased over time as it was used by periphytic communities (deduced from decreasing concentrations over time). Water was filtered through 0.45 μm membrane filters for DP and orthophosphate analysis. TP and DP samples were analyzed after persulfate digestion in an autoclave. Orthophosphate samples were pre-acidified with 0.01 mol/L HCl (ACS grade, Fisher Scientific, Hampton, New Hampshire, USA) (Carignan and Neiff 1992). Wash and standard solutions were used every 10 samples to ensure analytical quality.
Statistical analyses
For Chla and phosphorus results, concentrations for all treatments (Control, G1, G2, G3, P1, P2, and P3) and each sampling time (T0, T3, and T7) were compared using two-factor analysis of variance (two-way ANOVA). When required, Dunnett’s or Tukey’s post hoc tests were used for multiple comparisons. Multiple t tests were used for reactive and total dissolved phosphorus concentrations. Multiple comparisons were corrected using the Holm–Sidak method, computing adjusted p values. Prior to the analyses, normality and homoscedasticity were verified and data were log+1 transformed when these assumptions were not satisfied. Significance level was set at p < 0.05. GraphPad Prism 7 (GraphPad Software Inc., La Jolla, California, USA) was used for statistical analyses.
Principal coordinate analyses (PCoA) for 16S and 18S data from UniFrac distance matrices were calculated and visualized using the phyloseq (McMurdie and Holmes 2013) and ggplot2 (Wickham 2009) packages in R (R Core Team 2016). Sample clustering hypotheses were tested using a permutational multivariate analysis of variance (permanova, adonis()) in vegan (Oksanen et al. 2016), as this test has been proven to be more powerful in detecting differences in community structure even when group dispersions are heterogeneous (Anderson and Walsh 2013). Homogeneity of dispersion among sample groups was assessed using betadisper() in vegan.
Results
Glyphosate concentrations
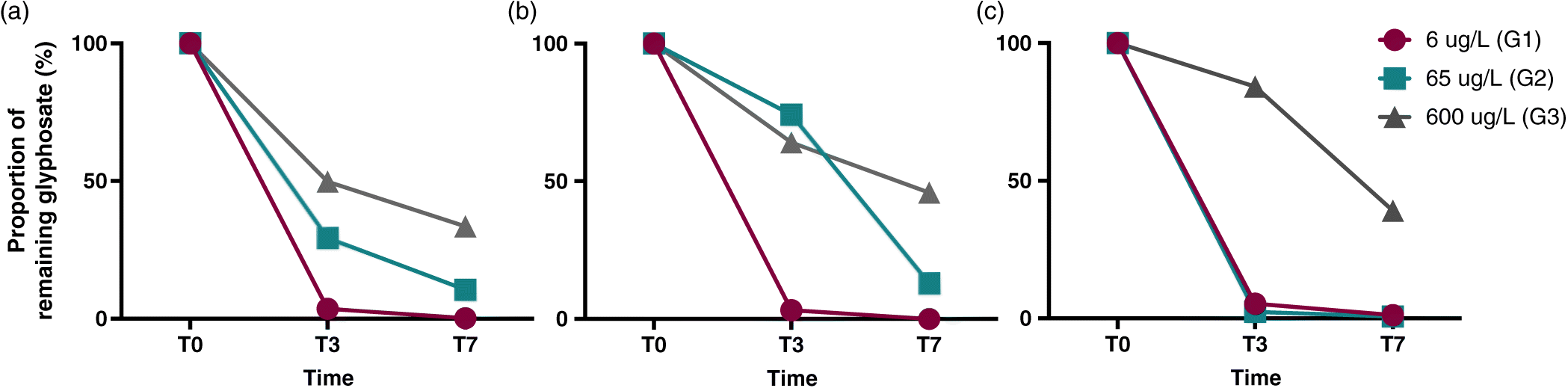
Biofilms of three different ages collected on artificial substrates were exposed to increasing concentrations of glyphosate. Our results showed that glyphosate concentrations in water decreased throughout all experiments by 55% to 100%, with no glyphosate remaining in the G1 series for all biofilm ages (Figs. 1a–1c). We also observed a slower relative rate of loss of glyphosate (average of 8.7% per day) at high concentrations (G3) compared with lower concentrations (G1) (average of 14.2% per day) (Figs. 1a–1c). For all three experiments, in the 65 (G2) and 600 μg/L (G3) treatments (Figs. 1a–1c), periphyton was exposed to the herbicide throughout the experiment. The same loss pattern was observed in a preliminary experiment we conducted in 2015 with 20-year-old biofilms under the same experimental conditions and for which absolute concentrations were measured (Fig. S4). This 2015 preliminary study also confirmed that our initial concentrations were accurate, as we repeated the same glyphosate additions in 2016.
Fig. 1.
Dissolved and reactive phosphorus
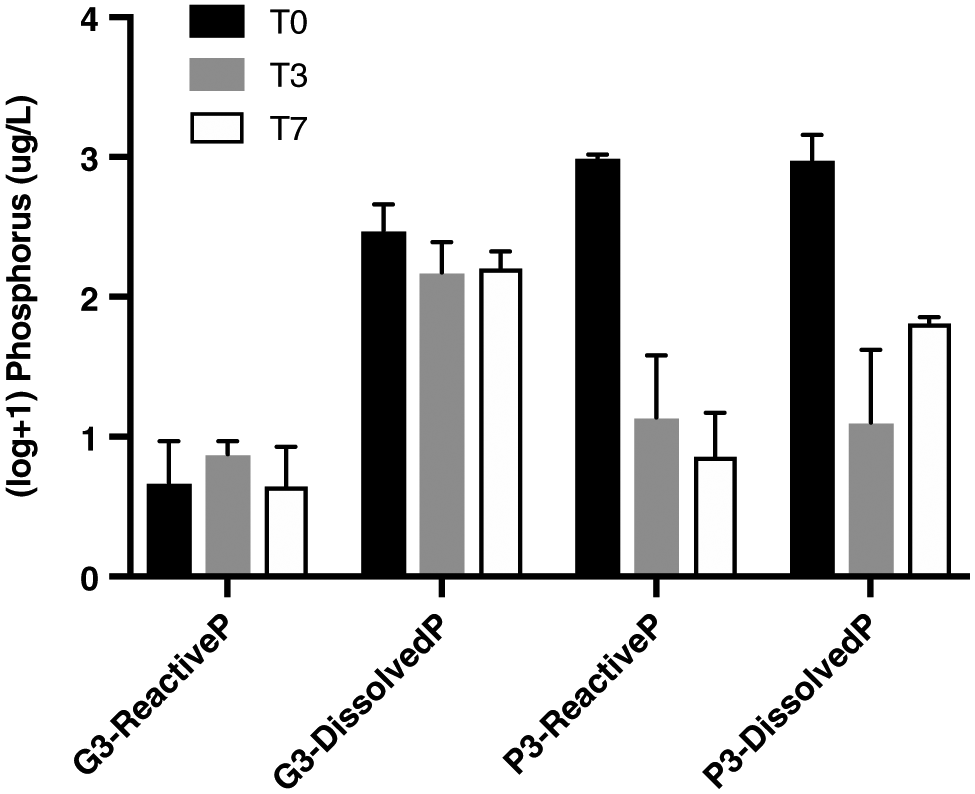
One of our main objectives was to compare the amount of phosphorus potentially available for algal growth in the glyphosate and phosphorus treatments. Only G3 and P3 treatments were plotted (Fig. 2) because orthophosphate concentrations were often low in the other treatments. The results showed that at the beginning of the experiment (T0), shortly after phosphorus was added, the amount of reactive phosphorus was about 300 times higher in the P3 treatment than in the G3 treatment. There were no significant changes in reactive phosphorus concentrations in the G3 treatment for all three experiments. However, the decrease in reactive phosphorus was significant in the P3 treatment between T0 and T7 (t test, padj < 0.0005). This was also the case in the P3 treatment between T0 and T7 for the 2-month-old biofilms (t test, padj < 0.01) and between T0 and T3 for the 1-year-old biofilms (t test, padj < 0.000005) (data not shown). Dissolved phosphorus concentrations did not vary significantly in the G3 treatment, but did in the P3 treatment, between T0 and T7 (t test, p < 0.0005). However, the herbicide appears to release a significant amount of total dissolved phosphorus, at concentrations similar to those measured at T0 in the P3 treatment.
Fig. 2.
Chlorophyll a
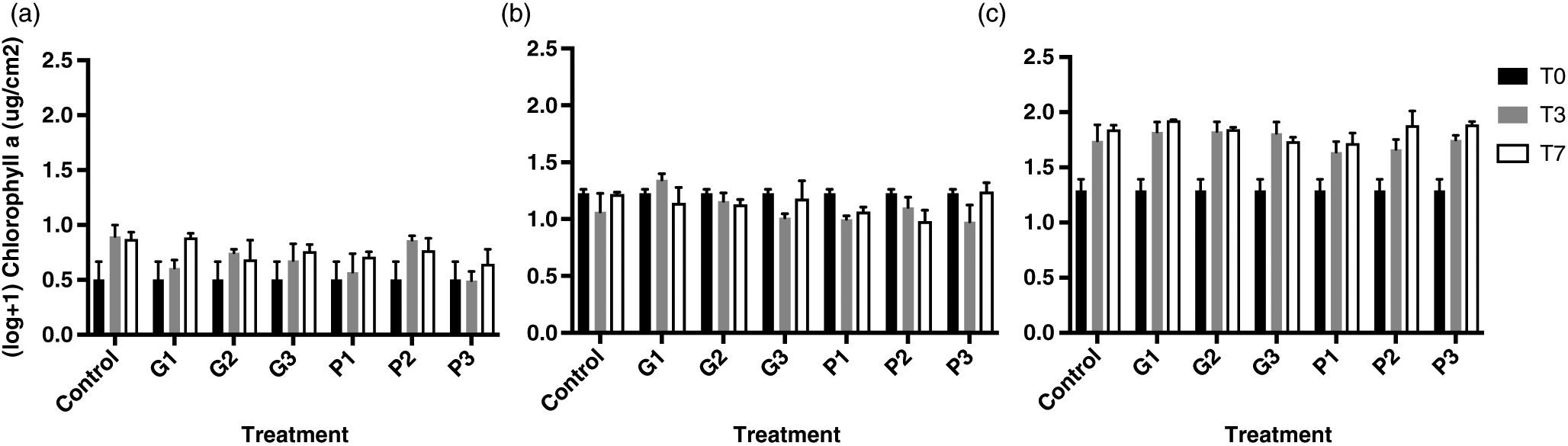
There was no significant decrease in Chla concentrations for all three submersion period experiments. The duration of exposure or treatment had no significant effect on Chla for 2-month-old (Fig. 3a) and 1-year-old periphyton (Fig. 3b) (p > 0.05) (multifactorial two-way ANOVA). As shown in Fig. 3c, the effect of time was significant for the oldest biofilms (two-way ANOVA, p < 0.0001). However, the effect of treatment was not significant and there was no interaction between treatment and time on Chla (p > 0.05). For all treatments (Fig. 3c), there was a significant increase between T0 and T3 and between T0 and T7 (Tukey’s honest significant difference following two-way ANOVA) (Table S1).
Fig. 3.
Community composition
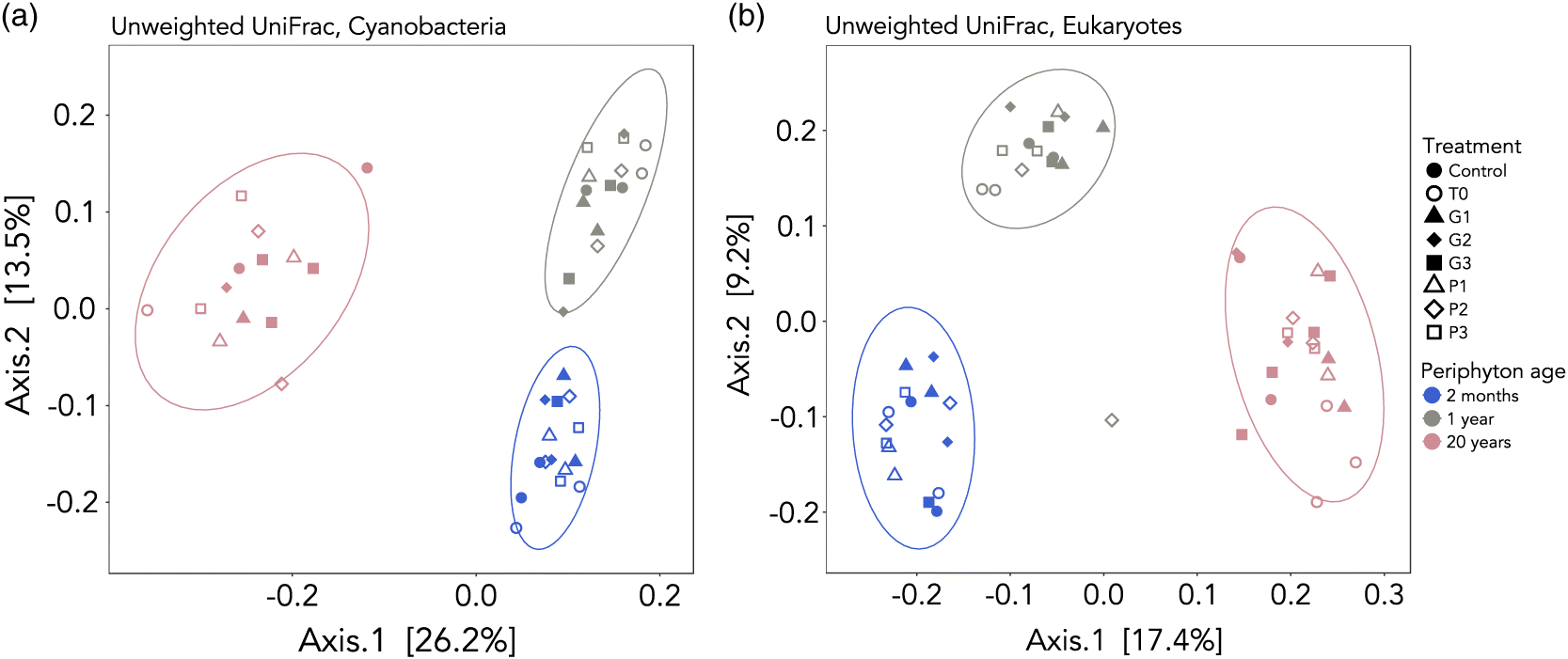
Using the unweighted distance metric UniFrac in beta diversity analyses, we found a significant clustering of samples by periphyton age, explaining 37% of the variation in community composition for cyanobacteria (adonis, p = 0.001) and 26% for eukaryotes (adonis, p = 0.001) (Fig. 4). Unweighted UniFrac is a presence/absence information analysis, rather than a relative abundance analysis (weighted UniFrac). We assessed both distance metrics, but cluster discrimination was sharper with unweighted UniFrac for eukaryotes (Fig. 4) (the results of the weighted UniFrac analyses are shown in Fig. S5). Multivariate dispersion was also significant for periphyton age (betadisper, p = 0.000525 and p = 0.0013 for cyanobacteria and eukaryotes, respectively), with greater dispersion within the 20-year-old samples for cyanobacteria and within the 2-month-old samples for eukaryotes. Treatment and sampling time did not significantly explain the variance in beta diversity.
Fig. 4.
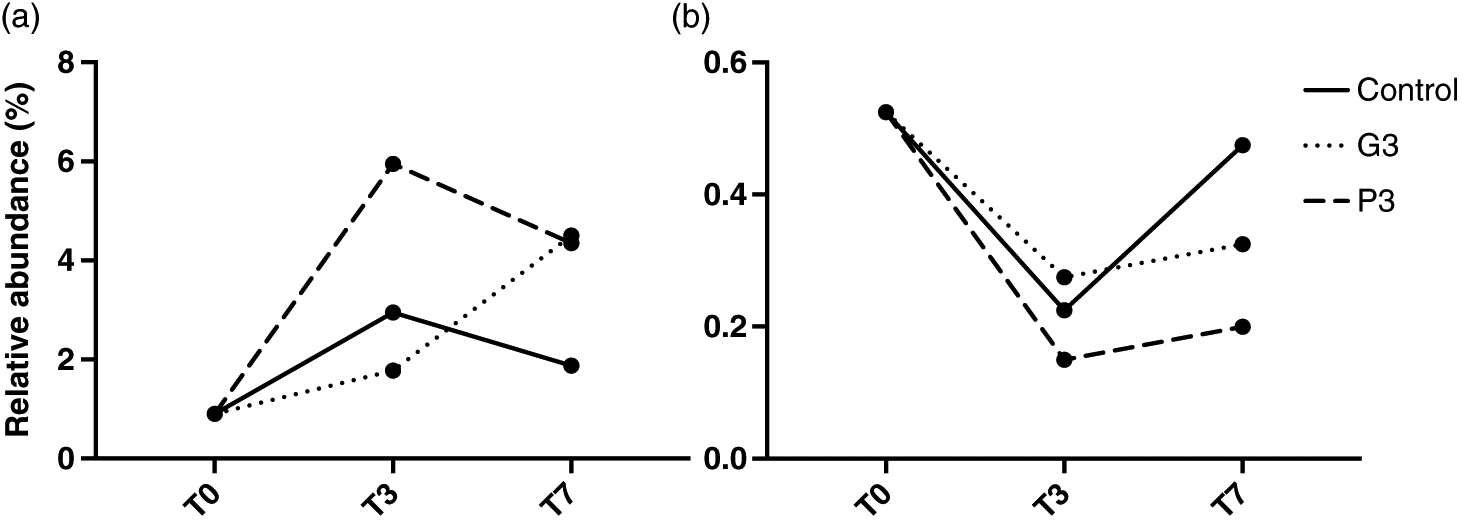
Following this community composition analysis, we assessed the relative abundance of the cyanobacterial genera Anabaena and Microcystis (Fig. 5) for control, G3, and P3 treatments throughout the experiment. This relative abundance was assessed as a fraction of the rarefied number of sequences/sample for 16S. Results show that, compared with the control and P3 treatments, the relative abundance of Anabaena increased in the G3 treatment between T0 and T7. In contrast, the relative abundance of Microcystis decreased between T0 and T3 for all three treatments and a slight increase was then observed between T3 and T7.
Fig. 5.
Discussion
Environmentally relevant glyphosate concentrations are lost from water within 7 d under our experimental conditions
Our results showed that local environmentally relevant concentrations of glyphosate degrade entirely within 7 d, diminishing the exposure of aquatic organisms to the herbicide. This contrasts with the work of Vera et al. (2010), who exposed periphytic communities to 8 mg/L of glyphosate and estimated a half-life of 4.2 d. This difference could be because of the fact that the latter experiment was conducted in large outdoor mesocosms with well-established bacterial communities in the water column. We attribute the observed degradation to the bacteria and fungi present in the biofilms, because glyphosate appears to be resistant to photodegradation (Aparicio et al. 2013). It is also possible that the microcosms’ filtered (0.45 μm) lake water still contained bacteria that could have contributed to glyphosate degradation. Preliminary results from our 2015 experiment showed that in microcosms where periphyton was exposed to 600 μg/L, AMPA was detected at low concentrations (6–7 μg/L) at T3 and T7 (Fig. S6), implying degradation. However, the loss of glyphosate that we observed could also be due to other mechanisms such as adsorption and (or) uptake by biofilms (Bengtsson et al. 2004; Evans-White and Lamberti 2009; Van den Brink et al. 2009). This uptake could be influenced by periphyton total biomass, which increases with submersion period, explaining differences in the loss of glyphosate in water between experiments. Even if glyphosate uptake was not directly measured, our results clearly showed that G1 and G2 initial concentrations decreased below our detection limit after 3 d in the 20-year-old biofilm experiment. Adsorption to the container walls could also explain glyphosate elimination from the water.
Our results also showed that after 7 d, biofilms from the G3 treatment were still exposed to relatively high concentrations of glyphosate. It is known that exposure of aquatic organisms to pesticides increases very rapidly following rainfall events and can reach relatively high concentrations during flood events (Tlili et al. 2008). Peruzzo et al. (2008) sampled water, sediments, and soil after glyphosate application and rain events in an agricultural area in Argentina. The highest glyphosate concentrations in the water samples were measured right after the first significant rain event, reaching a maximum of 700 μg/L. This level is comparable with our highest treatment, in which we measured the slowest relative rate of loss. Our experimental design, therefore, seems to accurately mimic short-term pulses following glyphosate application, suggesting that significant leaching or flooding due to heavy rains immediately following glyphosate spraying could potentially lead to significant exposure in aquatic biota. Glyphosate application frequency per season and the occurrence of heavy rain events, therefore, play a crucial role in understanding the consequences of pulse exposure on biological populations.
Glyphosate is a source of phosphorus but not in a bioavailable form
In aquatic ecosystems and soils, glyphosate is rapidly and completely degraded by microorganisms to water, carbon dioxide, and inorganic phosphate (Aparicio et al. 2013). Two main pathways were identified for glyphosate degradation, both resulting in the breakage of the molecule’s C–P bond. In the first pathway, the herbicide is degraded to AMPA and glyoxylate through the activity of a glyphosate oxidoreductase. AMPA is then either directly metabolized to methylamine and orthophosphate or acetylated prior to the cleavage of the C–P bond (Forlani et al. 2008). In the second pathway, this cleavage is caused by a C–P lyase complex, leading to the release of purines and amino acids (Aparicio et al. 2013). Although the amount of total phosphorus released by glyphosate through this C–P cleavage has been reported in the literature (Lipok et al. 2007; Vera et al. 2010; Vera et al. 2014; Gomes et al. 2016b), dissolved reactive phosphorus has never been considered. Our results showed that glyphosate releases a significant amount of phosphorus into surrounding waters, but that this phosphorus does not seem to be bioavailable to biofilms. In fact, the amount of reactive phosphorus that was released while glyphosate was lost from water was low and did not decrease over time, implying that it was not used by periphytic communities.
In aquatic ecosystems, cyanobacteria play a major role in both carbon and nitrogen cycling, being the only prokaryotes capable of oxygenic photosynthesis and having the ability to fix atmospheric N2. Some strains of cyanobacteria have also been identified to exhibit a natural tolerance to glyphosate (Powell et al. 1991). Forlani et al. (2008) studied the ability of six strains of cyanobacteria to use glyphosate as a sole source of phosphorus. They found that four strains (Anabaena, Leptolyngbya, Microcystis, and Nostoc) out of six were able to grow on a medium that contained the herbicide as the only phosphorus source. Because they did not observe any release of inorganic phosphorus, the hypothesis that these cyanobacteria were able to hydrolyze the glyphosate molecule C–P’s bond was rejected. We observed an increase in the relative abundance of the glyphosate-resistant strain of the cyanobacterial genus Anabaena in comparison with the control as well as a slight increase in Microcystis between T3 and T7. Because we were able to detect low amounts of AMPA (Fig. S6) in our 2015 experiment under the same conditions, as well as low amounts of orthophosphates in the current study, we hypothesize that the cyanobacteria of the biofilms were able to degrade glyphosate via the first pathway.
Microcystis and Anabaena are globally detected in toxic blooms (Yoshida et al. 2008; Bláha et al. 2009; Chalifour et al. 2016), raising concern regarding the use of glyphosate, as this herbicide favours the growth of certain taxa even in the absence of bioavailable phosphorus. Vera et al. (2010) added 8 mg/L glyphosate as Roundup® to outdoor mesocosms with periphyton. After 3 d and 8 d, they measured approximately 800 μg/L TP, as this concentration did not vary significantly between the two sampling times. The input of total phosphorus through the addition of glyphosate is, therefore, fairly substantial, as was also shown by our results (Fig. S7). Vera et al. (2010) concluded that glyphosate might promote eutrophication in aquatic systems, favouring potentially toxic cyanobacterial blooms. This contradicts our results, as we found that this phosphorus input is not available to biofilms. However, it is possible that cyanobacteria could, in fact, use this phosphorus as a nutrient source, but that their number was not great enough in our biofilms. This could partially explain the absence of significative decreases in reactive and dissolved phosphorus for the G3 treatment.
Chlorophyll a and essential aromatic amino acid contents are not adversely impacted by glyphosate in biofilms
Because of its short half-life, glyphosate is rapidly degraded to AMPA, which is known to alter the biosynthesis of chlorophyll (Gomes et al. 2016a). Furthermore, we hypothesized that young biofilms would be more sensitive to glyphosate than mature biofilms because in older biofilms, microscopic niches are created and nutrient exchanges with the water column decrease, thus offering protection from external stressors. However, we did not observe any significant decrease in Chla content for any of the three submersion period experiments. This absence of adverse effect was also observed by Lozano et al. (2018) who exposed 60-day-old periphytic biofilms to 3 mg/L of pure glyphosate in microcosm experiments. After a colonization period of 60 d, biofilms developed a sufficiently consolidated matrix, limiting contact between the organisms and glyphosate. This latter hypothesis is consistent with our results, as our youngest biofilms were also 60 days old. It has been established that the main degradation product of glyphosate, AMPA, can alter the biosynthesis of Chla in plants (Gomes et al. 2014; Gomes et al. 2016a). To our knowledge, the adverse effects of AMPA on periphyton have yet to be explored. Preliminary results from our 2015 experiment showed that in microcosms where periphyton was exposed to 600 μg/L, AMPA was detected at low concentrations (6–7 μg/L) at T3 and T7 (Fig. S6). However, Gomes et al. (2016a) observed an adverse effect on Chla metabolism in willow plants at AMPA concentrations >3 g/L. Vera et al. (2010) and Pérez et al. (2007) also observed a decrease in Chla concentrations in periphyton exposed to 6–12 mg/L of glyphosate. These concentrations are between 10 and 100 times higher than the environmental concentrations that we tested, which might explain the absence of any observed adverse effect on Chla. Moreover, these two studies assessed the effects of the commercial formulation Roundup® rather than the active ingredient, also contrasting with our experimental design. Indeed, it has been shown that polyethoxylated tallowamine (POEA), the surfactant typically used in Roundup®, is more toxic than glyphosate itself (Struger et al. 2008).
In contrast with what we expected, we observed a significative increase in Chla for the 20-year-old periphyton, which is consistent with the work by Vera et al. (2014), who observed an increase in Chla content in periphyton exposed to 3 mg/L glyphosate. Vera et al. (2014) tested both the active ingredient and a commercial formulation and did not detect any significant difference. Glyphosate triggered pathways for the synthesis of proteins and metabolites involved in stress response, favouring growth and Chla biosynthesis. According to Vera et al. (2014), it is also possible that the herbicide was used as a source of nutrients by some species of cyanobacteria, also stimulating growth, as shown by Forlani et al. (2008). Because the increase that we measured in the glyphosate treatments was also observed in the control, the experimental effect hypothesis cannot be excluded. However, the most significant increase we observed was between T0 and T7 in the G1 treatment (Table S1).
The principal mode of action of glyphosate is the inhibition of the synthesis of the enzyme EPSPS in plants, which is involved in the biosynthesis of the essential aromatic amino acids phenylalanine, tyrosine, and tryptophan (Gomes et al. 2014). Because this effect had never been assessed in periphytic biofilms, we measured these amino acid concentrations in 20-year-old biofilms exposed to glyphosate for 7 d in the preliminary experiment that we conducted in 2015 (SM.2). However, we did not detect any adverse effects of glyphosate on the metabolism of essential aromatic amino acids in biofilms (Fig. S8). We, therefore, infer that glyphosate might not impact the biosynthesis of amino acids at a community level, but future work should consider the effects on specific populations within the biofilms.
Microbial communities are more influenced by biofilm age than glyphosate exposure
We assessed the community composition of contrasting biofilm ages using specific primers that target cyanobacteria. Even at concentrations 100 times higher than local environmental levels, glyphosate did not significantly disrupt periphytic communities. Age, in fact, was the only factor that clearly shaped community structure, contradicting our hypothesis. However, we observed an increase in the relative abundance of Anabaena in the G3 treatment after 7 d compared with the control (Fig. 5). This taxon has been identified to possess a rare resistant form of the EPSPS enzyme, targeted by the mode of action of glyphosate (Powell et al. 1991; Forlani et al. 2008). In contrast, we found that the relative abundance of Microcystis decreased in the G3 treatment after 3 d but increased slightly between T3 and T7. Even if this taxon does not carry the resistant form of the EPSPS enzyme, Forlani et al. (2008) found that Microcystis was able to grow on a medium containing glyphosate as the only phosphorus source. Although our sequencing methods do not allow us to verify the presence of the rare, resistant form of EPSPS identified by Forlani et al. (2008), we think that the increased relative abundance of Anabaena after 7 d in the G3 treatment compared with the control shows that it is resistant to glyphosate and favoured by the release of dissolved reactive phosphorus. The absence of a drastic temporal turnover in the community composition of our biofilms may be because of the relatively short duration of our experiments and the relatively low glyphosate levels that we tested. However, this study provides an interesting snapshot of the biological processes related to the exposure of periphytic biofilms to environmental concentrations of glyphosate under pulse event conditions.
Few studies consider the maturation stage and, hence, the stability of biofilms when assessing the effect of chemical contamination. Ivorra et al. (2000) compared the effects of metal contamination on 2- and 6-week-old biofilms. Their hypothesis was that as biofilms develop, internal cycling of materials dominates over diffusion and exchanges with the water column. Moreover, as biofilms mature, EPS production increases, limiting the penetrations of toxicants. Ivorra et al. (2000) found that older biofilms (6 weeks old) were more resistant to metal contamination than younger biofilms (2 weeks old). Guasch et al. (2004) exposed 5-week-old and 6-week-old periphytic communities from an oligotrophic stream to copper for several weeks. Contrary to their predictions, there were no differences in the physiological responses of low and high biomass biofilms exposed to copper. These two studies were conducted in streams, where water velocity plays an important role in periphyton thickness and microarchitecture (Besemer et al. 2012; Battin et al. 2016), in contrast with lake biofilms. The more mature biofilms that were studied (6 weeks old) by Ivorra et al. (2000) and Guasch et al. (2004) were also significantly younger and, thus, likely thinner than the ones we exposed in our study (20 years old). Given these two major differences, it is difficult to extrapolate the results of these two studies to our own observations. We, therefore, hypothesize that, in lakes, even younger biofilms (2 months old) are characterized by a thick EPS layer that limits the penetration of glyphosate, thereby preventing its adverse effects.
Conclusion
Periphytic biofilms are key organisms in ecotoxicological studies, as they are at the base of food webs, are important biogeochemical cyclers in streams and lakes, and are often the first communities to be in direct contact with runoff. This study assessed the effects of technical grade glyphosate on contrasting colonization stages of lake biofilms. The results showed that at levels that are currently detected in eastern Canadian rivers, glyphosate poses little risk for periphyton under pulse event conditions. In fact, no adverse effect was observed on Chla content. The herbicide was also found to release phosphorus in the water, but not in a bioavailable form, limiting uptake by biofilms. Periphyton age was the only factor shaping the community composition of biofilms, suggesting that the EPS layer produced by algae and bacteria was too thick to allow the penetration of glyphosate, even for the younger communities. Age and thickness likely caused important inter-experimental bias, and it is, therefore, critical to consider the colonization stage of biofilms before interpreting ecotoxicological effects. However, we identified an increase in the relative abundance of Anabaena, which has been previously found to carry a rare resistant form of the EPSPS enzyme targeted by glyphosate. This highlights the fact that some effects might be detected at a population or taxon scale, but not at a community scale. Future studies should, thus, include specific assessment of glyphosate’s toxicity using biomarkers such as reactive oxygen species or enzymes. More attention should also be given to cyanobacteria, as toxic strains seem favoured by the use of the herbicide. As specific primers allowed us to monitor the presence and abundance of cyanobacteria, future studies should focus on elucidating their mechanism of resistance. In conclusion, this study represents a step towards understanding the effects of a non-selective herbicide of rising concern on lake ecosystems.
Acknowledgements
This project was financially supported by a FRQ-NT Programme d’initiatives stratégiques pour l’innovation (ISI) grant to M.A. and D.P, by an NSERC Discovery grant to M.A, and by the Canada Research Chair program to M.A. M.K. received a scholarship through the NSERC CREATE ÉcoLac training program in Lake and Fluvial Ecology. We acknowledge the Integrated Microbiome Resource (IMR) lab at Dalhousie University (Halifax, Canada) for the library preparation of 16S and 18S amplicons, sequencing, and standard bioinformatics analysis pipeline. We acknowledge the Metabolics Core Facility at McGill University (Quebec, Canada) for the amino acid analyses. We are grateful for laboratory assistance from Dominic Bélanger, Jérémy De Bonville, Antoine Caron, Marie-Christine Lafrenière, and Serge Paquet, and field assistance from Jérémy De Bonville and Maxime Leclerc. We acknowledge the staff of the Station de biologie des Laurentides (Université de Montréal) for logistical support.
References
Anderson MJ, and Walsh DCI. 2013. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: what null hypothesis are you testing? Ecological Monographs, 83(4): 557–574.
Andrews S. 2010. FastQC: a quality control tool for high throughput sequence data [online]: Available from bioinformatics.babraham.ac.uk/projects/fastqc/.
Aparicio VC, De Gerónimo E, Marino D, Primost J, Carriquiriborde P, and Costa JL. 2013. Environmental fate of glyphosate and aminomethylphosphonic acid in surface waters and soil of agricultural basins. Chemosphere, 93(9): 1866–1873.
Battin TJ, Besemer K, Bengtsson MM, Romani AM, and Packmann AI. 2016. The ecology and biogeochemistry of stream biofilms. Nature Reviews Microbiology, 14(4): 251–263.
Benbrook CM. 2016. Trends in glyphosate herbicide use in the United States and globally. Environmental Sciences Europe, 28(1): 3.
Bengtsson G, Hansson L-A, and Montenegro K. 2004. Reduced grazing rates in Daphnia pulex caused by contaminants: implications for trophic cascades. Environmental Toxicology and Chemistry, 23(11): 2641–2648.
Besemer K, Peter H, Logue JB, Langenheder S, Lindstrom ES, Tranvik LJ, and Battin TJ. 2012. Unraveling assembly of stream biofilm communities. The ISME Journal, 6(8): 1459–1468.
Bláha L, Babica P, and Maršálek B. 2009. Toxins produced in cyanobacterial water blooms—toxicity and risks. Interdisciplinary Toxicology, 2(2): 36–41.
Bregnard TP-A, Höhener P, Häner A, and Zeyer J. 1996. Degradation of weathered diesel fuel by microorganisms from a contaminated aquifer in aerobic and anaerobic microcosms. Environmental Toxicology and Chemistry, 15(3): 299–307.
Bushnell B. 2014. BBMap: a fast, accurate, splice-aware aligner [online]: Available from osti.gov/servlets/purl/1241166.
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. 2010. QIIME allows analysis of high-throughput community sequencing data. Nature Methods, 7(5): 335–336.
Carignan R, and Neiff JJ. 1992. Nutrient dynamics in the floodplain ponds of the Paraná River (Argentina) dominated by the water hyacinth Eichhornia crassipes. Biogeochemistry, 17: 85–121.
CEAEQ. 2008. Détermination du glyphosate et de l′AMPA dans les eaux: dosage par chromatographie en phase liquide; dérivation post-colonne et détection en fluorescence. MA. 403—GlyAmp 1.0, Rév. 3, Ministère du Développement durable, de l′Environnement et des Parcs du Québec, Québec City, Québec [online]: Available from ceaeq.gouv.qc.ca/methodes/pdf/MA403GlyAmp10.pdf.
Chalifour A, LeBlanc A, Sleno L, and Juneau P. 2016. Sensitivity of Scenedesmus obliquus and Microcystis aeruginosa to atrazine: effects of acclimation and mixed cultures, and their removal ability. Ecotoxicology, 25(10): 1822–1831.
Desrosiers M, Planas D, and Mucci A. 2006. Mercury methylation in the epilithon of boreal shield aquatic ecosystems. Environmental Science & Technology, 40(5): 1540–1546.
Edgar RC, Haas BJ, Clemente JC, Quince C, and Knight R. 2011. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics, 27(16): 2194–2200.
Evans-White MA, and Lamberti GA. 2009. Direct and indirect effects of a potential aquatic contaminant on grazer–algae interactions. Environmental Toxicology and Chemistry, 28(2): 418–426.
Forlani G, Pavan M, Gramek M, Kafarski P, and Lipok J. 2008. Biochemical bases for a widespread tolerance of cyanobacteria to the phosphonate herbicide glyphosate. Plant and Cell Physiology, 49(3): 443–456.
Gaffney AM, Markov SA, and Gunasekaran M. 2001. Utilization of cyanobacteria in photobioreactors for orthophosphate removal from water. Applied Biochemistry and Biotechnology, 91(1–9): 185–194.
Giroux I. 2015. Présence de pesticides dans l’eau au Québec: portrait et tendances dans les zones de maïs et de soya—2011 à 2014. ministère du Développement durable, de l’Environnement et de la lutte contre les changements climatiques, Direction du suivi de l’état de l’environnement, Québec City, Québec, 47 p. + 5 ann [online]: Available from mddelcc.gouv.qc.ca/pesticides/mais_soya/portrait2011-2014/index.htm.
Gomes MP, Smedbol E, Chalifour A, Hénault-Ethier L, Labrecque M, Lepage L, et al. 2014. Alteration of plant physiology by glyphosate and its by-product aminomethylphosphonic acid: an overview. Journal of Experimental Botany, 65(17): 4691–4703.
Gomes MP, Le Manac’h SG, Maccario S, Labrecque M, Lucotte M, and Juneau P. 2016a. Differential effects of glyphosate and aminomethylphosphonic acid (AMPA) on photosynthesis and chlorophyll metabolism in willow plants. Pesticide Biochemistry and Physiology, 130: 65–70.
Gomes MP, Le Manac’h SG, Moingt M, Smedbol E, Paquet S, Labrecque M, et al. 2016b. Impact of phosphate on glyphosate uptake and toxicity in willow. Journal of Hazardous Materials, 304: 269–279.
Gordon A. 2009. FASTX-Toolkit: FASTQ/A short-reads pre-processing tools [online]: Available from hannonlab.cshl.edu/fastx_toolkit/.
Guasch H, Navarro E, Serra A, and Sabater S. 2004. Phosphate limitation influences the sensitivity to copper in periphytic algae. Freshwater Biology, 49: 463–473.
Hagerthey SE, Bellinger BJ, Wheeler K, Gantar M, and Gaiser E. 2011. Everglades periphyton: a biogeochemical perspective. Critical Reviews in Environmental Science and Technology, 41(Suppl. 1): 309–343.
Hamelin S, Amyot M, Barkay T, Wang Y, and Planas D. 2011. Methanogens: principal methylators of mercury in lake periphyton. Environmental Science & Technology, 45(18): 7693–7700.
Ivorra N, Bremer S, Guasch H, Kraak MHS, and Admiraal W. 2000. Differences in the sensitivity of benthic microalgae to Zn and Cd regarding biofilm development and exposure history. Environmental Toxicology and Chemistry, 19(5): 1332–1339.
Lavoie I, Lavoie M, and Fortin C. 2012. A mine of information: benthic algal communities as biomonitors of metal contamination from abandoned tailings. Science of the Total Environment, 425: 231–241.
Lázaro WL, Guimarães JR, Ignácio AR, Da Silva CJ, and Díez S. 2013. Cyanobacteria enhance methylmercury production: a hypothesis tested in the periphyton of two lakes in the Pantanal floodplain, Brazil. Science of the Total Environment, 456–457: 231–238.
Leclerc M, Planas D, and Amyot M. 2015. Relationship between extracellular low-molecular-weight thiols and mercury species in natural lake periphytic biofilms. Environmental Science & Technology, 49(13): 7709–7716.
Lipok J, Owsiak T, Młynarz P, Forlani G, and Kafarski P. 2007. Phosphorus NMR as a tool to study mineralization of organophosphonates—the ability of Spirulina spp. to degrade glyphosate. Enzyme and Microbial Technology, 41(3): 286–291.
Lozano VL, Vinocur A, Sabio y García CA, Allende L, Cristos DS, Rojas D, et al. 2018. Effects of glyphosate and 2, 4-D mixture on freshwater phytoplankton and periphyton communities: a microcosms approach. Ecotoxicology and Environmental Safety, 148: 1010–1019.
Lozupone C, and Knight R. 2005. UniFrac: a new phylogenetic method for comparing microbial communities. Applied and Environmental Microbiology, 71(12): 8228–8235.
Lozupone C, Lladser ME, Knights D, Stombaugh J, and Knight R. 2011. UniFrac: an effective distance metric for microbial community comparison. The ISME Journal, 5(2): 169–172.
McMurdie PJ, and Holmes S. 2013. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE, 8(4): e61217.
Montuelle B, Dorigo U, Bérard A, Volat B, Bouchez A, Tlili A, et al. 2010. The periphyton as a multimetric bioindicator for assessing the impact of land use on rivers: an overview of the Ardières-Morcille experimental watershed (France). Hydrobiologia, 657(1): 123–141.
Nusch EA. 1980. Comparison of different methods for chlorophyll and phaeopigment determination. Archiv für Hydrobiologie—Beiheft Ergebnisse der Limnologie, 14: 14–36.
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, Mcglinn D, et al. 2016. vegan: community ecology package [online]: Available from cran.r-project.org/package=vegan.
Pérez GL, Torremorell A, Mugni H, Rodríguez P, Vera MS, Naschimento MD, et al. 2007. Effects of the herbicide Roundup on freshwater microbial communities: a mesocosm study. Ecological Applications, 17(8): 2310–2322.
Perron T, Chételat J, Gunn J, Beisner BE, and Amyot M. 2014. Effects of experimental thermocline and oxycline deepening on methylmercury bioaccumulation in a Canadian Shield lake. Environmental Science & Technology, 48(5): 2626–2634.
Peruzzo PJ, Porta AA, and Ronco AE. 2008. Levels of glyphosate in surface waters, sediments and soils associated with direct sowing soybean cultivation in north pampasic region of Argentina. Environmental Pollution, 156(1): 61–66.
Powell HA, Kerby NW, and Rowell P. 1991. Natural tolerance of cyanobacteria to the herbicide glyphosate. New Phytologist, 119: 421–426.
R Core Team. 2016. R: a language and environment for statistical computing [online]: Available from r-project.org/.
Rognes T, Flouri T, Nichols B, Quince C, and Mahé F. 2016. VSEARCH: a versatile open source tool for metagenomics. PeerJ, 4: e2584.
Schmitt-Jansen M, and Altenburger R. 2005. Toxic effects of isoproturon on periphyton communities—A microcosm study. Estuarine, Coastal and Shelf Science, 62(3): 539–545.
Smedbol E, Gomes MP, Paquet S, Labrecque M, Lepage L, Lucotte M, and Juneau P. 2018. Effects of low concentrations of glyphosate-based herbicide factor 540® on an agricultural stream freshwater phytoplankton community. Chemosphere, 192: 133–141.
Stevenson RJ, Bothwell ML, and Lowe RL. 1996. Algal ecology: freshwater benthic ecosystems. Academic Press, San Diego, California. 753 p.
Strickland JDH, and Parsons TR. 1972. A practical handbook of seawater analysis. 2nd ed. Fisheries Research Board of Canada, Ottawa, Ontario. 310 p.
Struger J, Thompson D, Staznik B, Martin P, McDaniel T, and Marvin C. 2008. Occurrence of glyphosate in surface waters of southern Ontario. Bulletin of Environmental Contamination and Toxicology, 80(4): 378–384.
Tlili A, Dorigo U, Montuelle B, Margoum C, Carluer N, Gouy V, et al. 2008. Responses of chronically contaminated biofilms to short pulses of diuron: An experimental study simulating flooding events in a small river. Aquatic Toxicology, 87(4): 252–263.
Tlili A, Bérard A, Roulier JL, Volat B, and Montuelle B. 2010. PO43− dependence of the tolerance of autotrophic and heterotrophic biofilm communities to copper and diuron. Aquatic Toxicology, 98(2): 165–177.
Tlili A, Corcoll N, Bonet B, Morin S, Montuelle B, Berard A, et al. 2011. In situ spatio-temporal changes in pollution-induced community tolerance to zinc in autotrophic and heterotrophic biofilm communities. Ecotoxicology, 20(8): 1823–1839.
Vadeboncoeur Y, and Steinman AD. 2002. Periphyton function in lake ecosystems. The Scientific World Journal, 2: 1449–1468.
Van den Brink PJ, Crum SJH, Gylstra R, Bransen F, Cuppen JGM, and Brock TCM. 2009. Effects of a herbicide–insecticide mixture in freshwater microcosms: risk assessment and ecological effect chain. Environmental Pollution, 157(1): 237–249.
Vera MS, Lagomarsino L, Sylvester M, Pérez GL, Rodriguez P, Mugni H, et al. 2010. New evidences of Roundup® (glyphosate formulation) impact on the periphyton community and the water quality of freshwater ecosystems. Ecotoxicology, 19(4): 710–721.
Vera MS, Juárez A, and Pizarro HN. 2014. Comparative effects of technical-grade and a commercial formulation of glyphosate on the pigment content of periphytic algae. Bulletin of Environmental Contamination and Toxicology, 93(4): 399–404.
Wetzel RG. 1983. Periphyton of freshwater ecosystems. Dr. W. Junk Publishers, The Hague, the Netherlands. 346 p.
Wetzel RG. 1993. Microcommunities and microgradients: linking nutrient regeneration, microbial mutualism, and high sustained aquatic primary production. Netherland Journal of Aquatic Ecology, 27(1): 3–9.
Wickham H. 2009. ggplot2: elegant graphics for data analysis. 1st ed. Springer-Verlag, New York City, New York. 216 p.
Yoshida M, Yoshida T, Kashima A, Takashima Y, Hosoda N, Nagasaki K, et al. 2008. Ecological dynamics of the toxic bloom-forming cyanobacterium Microcystis aeruginosa and its cyanophages in freshwater. Applied Environmental Microbiology, 74(10): 3269–3273.
Supplementary Material
Supplementary Material 1 (DOCX / 1.24 MB)
- Download
- 1.25 MB
Information & Authors
Information
Published In

FACETS
Volume 3 • Number 1 • October 2018
Pages: 934 - 951
Editor: Karen Kidd
History
Received: 5 June 2018
Accepted: 27 August 2018
Version of record online: 1 November 2018
Copyright
© 2018 Khadra et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
All relevant data are within the paper, the Supplementary Material, and the Sequence Read Archive (SRA) database (National Center for Biotechnology Information (NCBI)) (ncbi.nlm.nih.gov/sra; SRA accession SRP150239, BioProject PRJNA475256).
Key Words
Sections
Subjects
Plain Language Summary
Age matters: Submersion time shapes community composition in biofilm exposed to the herbicide glyphosate
Authors
Author Contributions
MK, DP, and MA conceived and designed the study.
MK performed the experiments/collected the data.
All analyzed and interpreted the data.
DP and MA contributed resources.
All drafted or revised the manuscript.
Competing Interests
The authors have declared that no competing interests exist.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Melissa Khadra, Dolors Planas, Catherine Girard, and Marc Amyot. 2018. Age matters: Submersion period shapes community composition of lake biofilms under glyphosate stress. FACETS.
3(1): 934-951. https://doi.org/10.1139/facets-2018-0019
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
Cited by
1. New insights into pesticide occurrence and multicompartmental monitoring strategies in stream ecosystems using periphyton and suspended sediment
2. Twenty Years of Research in Ecosystem Functions in Aquatic Microbial Ecotoxicology
3. How do metals interact with periphytic biofilms?
4. Microbiomes and glyphosate biodegradation in edaphic and aquatic environments: recent issues and trends
5. First evaluation of the periphyton recovery after glyphosate exposure
6. Resistance, resilience, and functional redundancy of freshwater bacterioplankton communities facing a gradient of agricultural stressors in a mesocosm experiment
7. Glyphosate-based herbicide exposure affects diatom community development in natural biofilms
8. Microbial Diversity and Mercury Methylation Activity in Periphytic Biofilms at a Run-of-River Hydroelectric Dam and Constructed Wetlands
9. Ecotoxicology of Glyphosate, Its Formulants, and Environmental Degradation Products
10. Community rescue in experimental phytoplankton communities facing severe herbicide pollution
11. Substratum‐associated microbiota