Incorporating protected areas into global fish biomass projections under climate change
Abstract
The world has set ambitious goals to protect marine biodiversity and improve ocean health in the face of anthropogenic threats. Yet, the efficiency of spatial tools such as marine reserves to protect biodiversity is threatened as climate change shifts species distributions globally. Here, we investigate the ability of global marine reserves to protect fish biomass under future climate change scenarios. Moreover, we explore regional patterns and compare worlds with and without marine reserves. We rely on computer modeling to simulate an utopian world where all marine reserves thrive and ocean governance is effective. Results suggest that climate change will affect fish biomass in most marine reserves and their surrounding waters throughout the 21st century. The biomass change varies among regions, with tropical reserves losing biomass, temperate ones gaining, and polar reserves having mixed effects. Overall, a world with marine reserves will still be better off in terms of fish biomass than a world without marine reserves. Our study highlights the need to promote climate resilient conservation methods if we are to maintain and recover biodiversity in the ocean under a changing world.
Introduction
The United Nations Convention on Biological Diversity (CBD) is discussing protecting 30% of the world’s oceans by 2030 (CBD 2021). Marine reserves are among the most popular tools for conservation (Song et al. 2017) and the main tool to reach the proposed target. No-take marine reserves (Costello and Ballantine 2015) can lead to increases in population biomass, density, resilience, individual size, and species diversity within the reserve’s boundaries (Micheli et al. 2004; Halpern 2014; Chirico et al. 2017; Friedlander et al. 2017). Such effects can lead to spillover of adult fish biomass in waters up to 2 km away, sometimes increasing fisheries yield (Halpern et al. 2009). While, in some cases, reserves have proven useful for promoting and protecting biomass (Halpern et al. 2009), improper implementation has led to little to no impact (Halpern 2003). Indeed, marine reserve performance both inside and outside of protected areas is not always clear (Hilborn et al. 2004; Ovando et al. 2021) and other socioecological variables are needed to maximize the benefits of marine reserves (Edgar et al. 2014; Grorud-Colvert et al. 2021). Moreover, as climate change shifts the distribution of marine species worldwide, questions remain regarding the impacts of such shifts to the effectiveness of marine reserves (Bruno et al. 2018; IPBES 2019; Jacquemont et al. 2022).
Climate change is one of the main threats to sustainable global ocean governance. The ocean has absorbed 90% of the excess heat produced by greenhouse gasses and captured about 30% of carbon emissions since the industrial revolution leading to warmer, more acidic, and less oxygenated oceans (IPCC 2022). This pattern is expected to continue through the 21st century. For example, sea surface temperature increase and dissolved oxygen decrease will have exceeded natural variability in about 40% of global protected areas by the mid-21st century (Bruno et al. 2018). Marine species have adopted different strategies to cope with these changes, with many shifting their known distribution by about 72 km per decade following the environmental conditions favorable to their development (Poloczanska et al. 2016). Moreover, modeling exercises suggest that such shifts are projected to continue regardless of the climate change scenario (Tittensor et al. 2021). The movement of species directly threatens conservation outcomes as most marine reserves are not designed with shifting species ranges in mind (Fredston-Hermann et al. 2018; Cashion et al. 2020).
The ecological effects of climate change are projected to be more intense at equatorial latitudes (Gattuso et al. 2015), a region where most marine reserves have been implemented (McLeod et al. 2009; Davies et al. 2017; UNEP-WCMC 2019). Climate change impacts could have a cascading effect on socioecological systems, such as fishing and ecotourism, that depend on marine reserves and their benefits from biological spillover. These impacts could also be exacerbated by other human activities such as coastal runoff (Crain et al. 2009), illegal fishing (Dalton et al. 2015; Dureuil et al. 2018), and water pollution (Schiff et al. 2011), ultimately threatening the conservation goals of marine reserves. Given the threats of climate change on marine species and the use of reserves as a common tool for achieving international goals, it is essential that we understand the relationship between marine reserves and climate change (IPBES 2019).
This study intends to contribute to the understanding of the hypothetical benefits of marine reserves in a changing world. Here, we ask (i) how is the biomass of marine reserves and its surrounding waters impacted by climate change? (ii) is that impact similar across different ecological regions? and (iii) would a changing world with marine reserves be better off in terms of fish biomass than a world without the protection of the marine reserves? To answer these questions, we combine a species distribution model with three Earth system models (ESMs) under two climate change and conservation scenarios. Our study is a simulation of a hypothetical world where all reserves are indeed fully no-take, well managed, and effective in promoting biomass. Moreover, we assume that marine reserves do not displace fishing effort to surrounding regions and fisheries are well managed outside protected waters. While such assumptions leads to a more utopian, rather than realistic, view of the current situation of the world’s marine reserves and fisheries, our simulation can help shed light on whether reserves maintain their benefits in terms of fish biomass in a changing world.
Methods
Accounting for marine reserves and species selection
We used the United Nations Environment World Conservation Monitoring Center’s World Database on Protected Areas (WDPA) as of December 2018 to determine the marine reserves in this study (UNEP-WCMC 2019). We define “marine reserve” as the marine protected areas (MPAs) labeled as “partial” or “full” no-take by the WDPA. The WDPA defines no-take as the complete halt of extracting any living or dead natural resources, with all methods of fishing, extraction, dumping, dredging, and construction, strictly prohibited in all or part of a protected area (UNEP-WCMC 2019). In total, we selected 596 no-take oceanic and coastal polygons (e.g., protected areas that consisted of only points were excluded; Table S1). In our simulation, all MPAs with any amount of no-take area were considered no-take, assuming that this could be a first step toward achieving full protection of 30% of the ocean by 2030. We gridded the WDPA onto a 0.5° latitude by 0.5° longitude map to harmonize the dataset with the spatial structure of our species distribution model (see the Projecting marine fish biomass under climate change section below). Given the ongoing discussions regarding marine reserves’ efficiency in both protecting biomass within and outside protected waters, we classified the grid cells into protected, surrounding, and unprotected. Protected waters include cells that contain a marine reserve polygon (cells that fully or partially contain a reserve from the WDPA) while cells that were immediately adjacent to a reserve cell were classified as surrounding waters. All other ocean cells were considered unprotected (Fig. S1). This allowed us to test the impacts of climate change to both protected and surrounding waters (see the Climate change impacts on marine reserves section below). The spatial designation analysis was conducted in ArcMap 10.6.1. To adjust for those reserve’s smaller than our protected cells, we estimated the proportion of each grid cell that contained a reserve using the “tabulate intersection” tool in ArcMap 10.6.1 and adjusted our estimates of fishing mortality by the proportion of the grid cell covered by the reserve. While we label these cells as “protected” for statistical comparisons, this proportional adjustment in fishing mortality accounts for the fact that many reserves are smaller than the grid cells used; thus, the “protected” fraction of the grid cell could go from 100% when the reserve is larger than the grid to <1% in the opposite case (see supplements – Accounting for reserve in fish biomass for more details). In this exercise, all reserves are assumed to be fully no-take, well managed, and effective in promoting biomass. We projected the biomass of 963 commercially important marine species under climate change for the whole globe. These species were selected for their life history data availability in FishBase (fishbase.org) and SeaLifeBase (sealifebase.org) as well as their importance for commercial fisheries (they cover 47% of the taxa included in the Sea Around Us catch database, representing 97% of the catch identified at the species’ level, and 56% of the world catch (e.g., including categories such as “Miscellaneous fin fishes”) between 2010 and 2014) (Zeller et al. 2016).
Projecting marine fish biomass under climate change
We used an established dynamic bioclimate envelope model (hereafter, DBEM) to project marine species biomass under different climate change scenarios (Cheung et al. 2016; Cheung et al. 2009a; Cheung et al. 2009b). Overall, the DBEM uses species depth limits, northern and southern latitudinal range limits, and species’ association with major habitat types to predict current distribution, and by considering species’ ecophysiology, growth and spatial population dynamics to project future changes in biomass and production under changing ocean conditions (Fig. 1.1). In each grid cell, biomass is estimated based on larvae and adult migration between cells, fishing, and intrinsic population growth parameters. For the conservation scenario, when the spatial cell is covered by a marine reserve, fishing mortality in that cell is proportional to the area covered by the reserve (see supplements – Accounting for reserve in fish biomass for more information and sensitivity analysis; Fig. S2). Larval dispersal from one cell to another is driven by ocean currents and diffusion, assuming that the recruitment magnitude is related to the larval duration in the water column (O’Connor et al. 2007), strength and direction of water currents, and diffusivity (Cheung et al. 2016). Mechanically speaking in the DBEM, as a grid cell reaches carrying capacity (e.g., protected grid cell) adults migrate to the neighboring cells (e.g., surrounding grid cells). Thus, in theory, waters protected from fisheries should have higher biomass than unprotected ones, consequently supplying biomass to unprotected cells, a process also known as spillover (Halpern et al. 2009). In our simulation, fisheries are well managed outside of protected waters, and the DBEM projects maximum catch potential (MCP) as a proxy of maximum sustainable yield for surrounding and unprotected waters. Both biomass and MCP of the modeled species are modeled at a 0.5° latitude by 0.5° longitude scale from 1951 to 2100 (see supplements and Cheung, Jones, et al. 2016; Cheung et al. 2009a; Cheung et al. 2009b for specific model details). While species distribution models are a useful tool in biogeography, their main limitation is that they do not include species interactions, potential evolutionary adaptation, and are subjected to the original data from which the model distribution was made (Cheung et al. 2009a).
We simulated biomass for each species using the climate forcings of three ESMs: the Geophysical Fluid Dynamics Laboratory Earth System Model 2M (GFDL, gfdl.noaa.gov/earth-system-model/), the Institute Pierre Simon Laplace Climate Model 5 (IPSL-CM5, cmc.ipsl.fr/ipsl-climate-models/ipsl-cm5/), and the Max Planck Institute for Meteorology Earth System Model (MPI, mpimet.mpg.de/en/science/models/) (Fig. 1.3). The original 1° × 1° grid cell of each ESM was downscaled to match the DBEM grid of 0.5° × 0.5° using the nearest neighbor method, and in some cases, bilinear interpolation (Cheung, Jones, et al. 2016). Results were averaged by time periods representing the present time (1995–2014), the mid-21st century (2041–2060), and the end of the 21st century (2081–2100) to reduce climate variability. We included two climate change scenarios (Fig. 1.1) following two Representative Concentration Pathways (RCPs) set by the Intergovernmental Panel on Climate Change (Meinshausen et al. 2011): a low greenhouse gas emission (strong mitigation, RCP 2.6) and a high emission (no mitigation, extreme, RCP 8.5) scenario (Riahi et al. 2011; van Vuuren et al. 2011). These RCPs represent contrasting climate change scenarios capturing the main spectrum of possible societal actions regarding mitigation used by the IPCC (IPCC 2022). Results are presented as the average of the ESMs to capture the structural uncertainty between models.
Climate change impacts on marine reserves
To explore the impacts of climate change in marine reserves (question i), we evaluated the percentage change in biomass between three time periods (change in time (ΔT) – Fig. 1.5; present, mid-century, and end of the 21st century) as follows (Fig. 1.5):where Bx is the biomass at mid or end of the 21st century and By is the biomass at present time. As an arbitrary rule, for all cases where Bx = 0 but By > 0, ΔT = −100, contrarily, if Bx > 0 but By = 0, ΔT = 100, and vice versa, naturally if both Bx and By = 0, then ΔT = 0. These rules applied to less than 4% of our grid for any time step, model, or RCP, but at the end of the century under RCP 8.5 (Supplementary Table S2).
We performed a Chi-squared test to answer (ii) whether the impacts of climate change would be similar across different regions. This allowed us to explore significant differences in the direction by which biomass would change (positive or negative) through the 21st century, relative to today. For this, we first categorized each grid cell according to the biome (e.g., template, tropical, polar) that each marine reserve was located in (Reygondeau 2019). We separated the temperate biome into northern and southern categories and only included polar south as our database has no reserves in the polar north biome. Since ΔT is a percentage value, we counted the amount of protected and non-protected grid cells that were projected to decrease in ΔT (e.g., ΔT < 0) and those that would increase or stay the same (e.g., ΔT >= 0) by the mid and end of the 21st century, relative to present time. Grid cells were tested by category (e.g., protected or surrounding) and biome separately (e.g., one test for grid cells containing a marine reserve in tropical biome and one test for surrounding in the same biome) at each future time step.
To explore differences between conservation scenarios (question iii), we ran a version of the DBEM where we did not incorporate marine reserves (Fig. 1.4, top) and fishing was possible in all regions (Cheung et al. 2009b). We first estimated the biomass difference between the two models in each time period. If the biomass difference was the same or decreased (e.g., 10% biomass difference between conservation scenarios in all time periods, or 15% difference today vs 10% by the mid-21st century), then marine reserves have no (when the same) or negative (when reducing) effect under climate change. On the other hand, if such difference increases over time (e.g., a 15% difference today vs a 20% by the mid-21st century), then marine reserves had positive effects on biomass even under climate change. The premise surrounding this analysis is that based on our methodology, the conservation model yields more biomass, and thus, such difference would only be impacted by climate change effects as other variables remain constant (note that comparing the raw biomass between models would be misleading, for the same reason). We also estimated ΔT for the no-conservation model (i.e., previous equation) and estimated the difference between Δ’s by subtracting the ΔT from the DBEM with marine reserves from the ΔT of the DBEM without them, that is, ΔB = ΔT conservation − ΔT no-conservation (ΔB; Fig. 1.6–7). Thus, a positive ΔB meant that marine reserves had a positive effect on buffering the impacts of climate change.
All of the data analysis and statistics were completed with the statistical software R version 3.6.2 (2019-12-12) and can be found at github.com/jepa/climatechangempa. Data from both the conservation and no-conservation models were not normally distributed, presenting high skewness and a sharper peak (leptokurtic according to the kurtosis test done with the moments package in R (Komsta and Novomestky 2015)). We explored the potential spatial autocorrelation of grid cells by computing the data semivariance (e.g., semi-variogram) using the R package nlme (Pinheiro and Bates 2020) (see supplemental material; Fig. S3). Statistical analyses were focused on protected and surrounding cells (i.e., the grid cells immediately adjacent to a protected cell). It is important to mention that the objective (and design) of this study was not to test the plausibility of protected waters supplying biomass to surrounding waters (i.e., spillover effect). We acknowledge that in the real world, spillover effect is case specific (Hilborn and Sinclair 2021; Ovando et al. 2021) and, when reported, often smaller (in terms of spatial scale) than our grid (Halpern et al. 2009). While our hypothetical world assumes spillover effect happens in all reserves (as a mechanistic part of the DBEM), we limit this effect to only the surrounding grid cells of a protected cell. All unprotected grid cells were located > 50 km from any protected grid cell (100 km when corrected for spatial autocorrelation), thus attributing the effects of the marine reserve for all of our species in these cells would be ecologically unsound. Our results should not be interpreted as evidence of a spillover effect on global marine reserves but rather an evaluation of the effect’s hypothetical performance under climate change.
Results
To answer our first question (i) how is the biomass of marine reserves and its surrounding waters impacted by climate change?, we compared the projected biomass of the conservation model in the mid and end of the 21st century relative to the present time (Fig. 1.4 bottom). As expected, global marine ecosystems will be impacted differently by climate change due to their natural ecosystemic structures and processes. Our results mirror these patterns by presenting an infinitely small average change in global biomass relative to today, yet large regional variations.
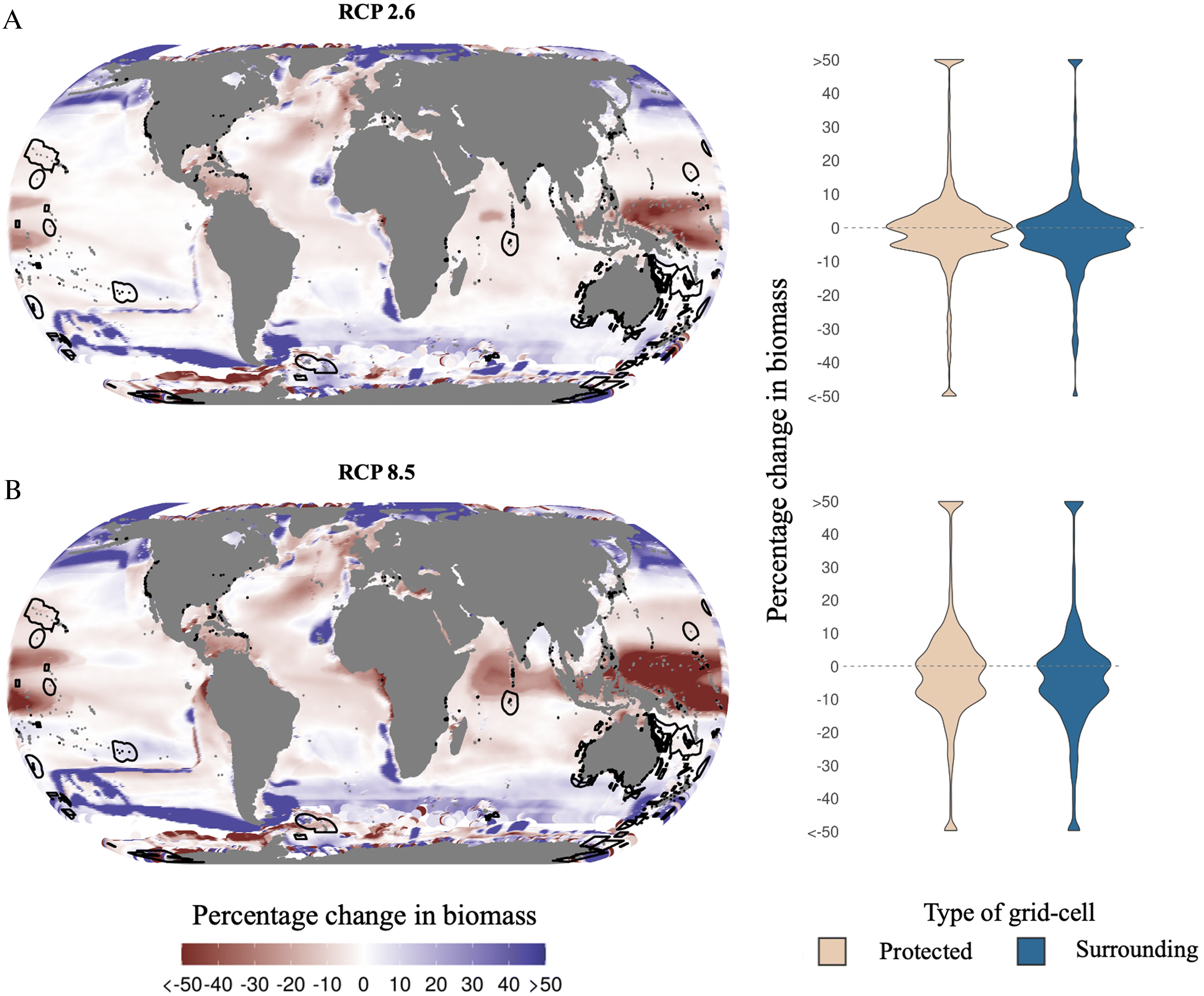
By the mid-21st century, marine reserves in our study are projected to have a global average change in biomass relative to today of −1% and 0% under a low and high emission scenario, respectively (Fig. 2). However, such a small (global) value is masked by a large variation (s.d.) of results ranging from ± 24% to ± 28% in a low and high emission scenario, respectively (Fig. 2B). Indeed major changes in biomass (i.e., > ± 50% change) are expected in some cases, regardless of the climate change scenario (Fig. 2). A similar pattern can be seen in surrounding waters where global change averages from 0% (s.d. 17%) under a low emission scenario to −2% (s.d. 25%) under a high emission scenario (Fig. 2B). Toward the end of the 21st century, changes in biomass will intensify under the high emission scenario while fewer extreme changes are expected under the low emission climate change scenario (Fig. S4). By then, average changes in biomass within protected waters will range from 3% (s.d. 26%) under a low emission scenario to 0% (s.d. 45%) under a high emission scenario while surrounding waters will change between 0% (s.d. 19%) to −12% (s.d. 46%) biomass under a low and high emission scenario, respectively. Note that variation (s.d.) by the end of the 21st century is larger, suggesting more extreme positive and negative impacts.
Fig. 1.
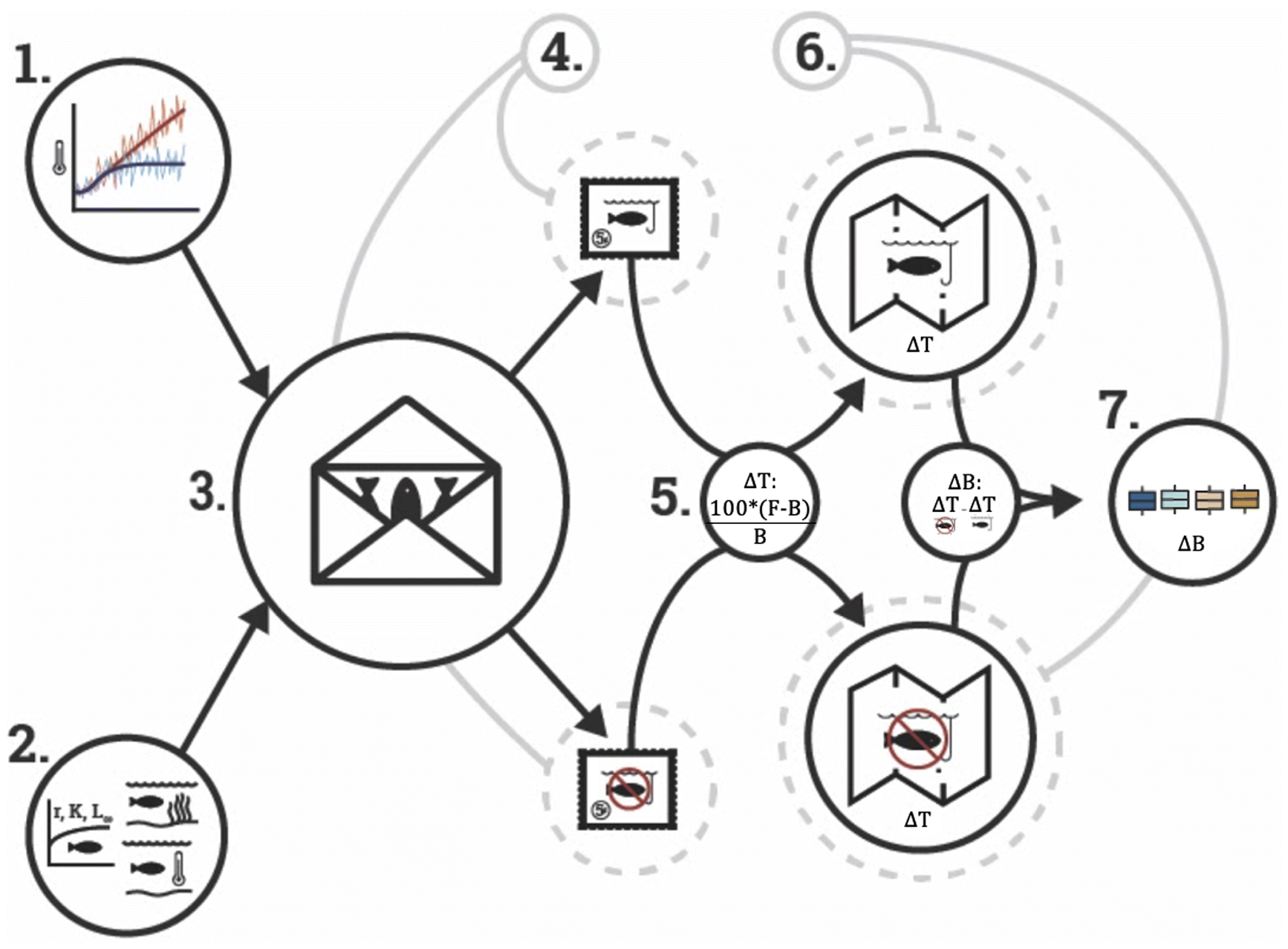
Fig. 2.
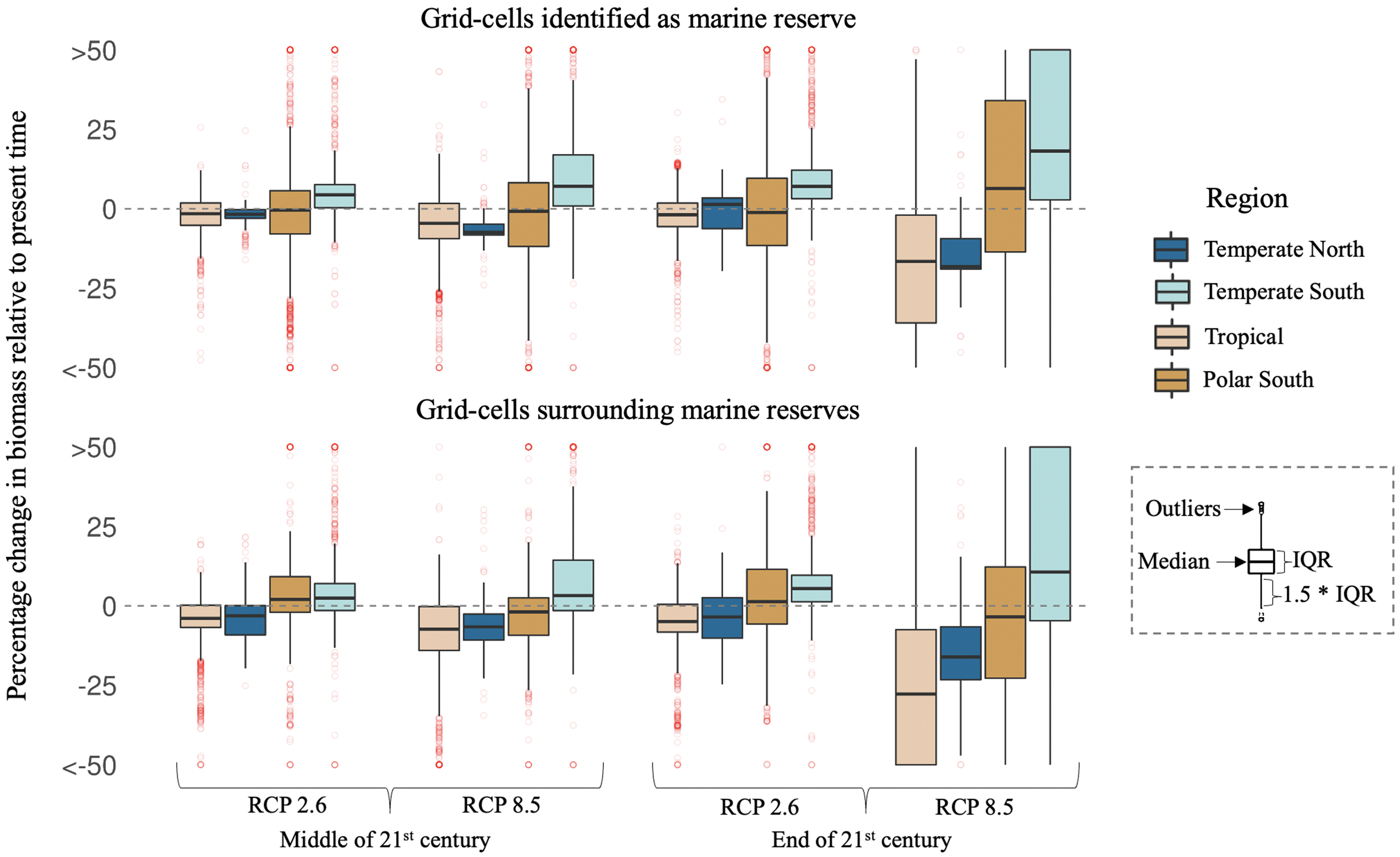
We ran a Chi-squared goodness of fit test in the conservation scenario to explore regional differences in the projected direction of change in biomass (e.g., positive or negative change) by the mid and end of the 21st century, relative to today (question ii). Overall, the direction of biomass change was not equally distributed (p < 0.05) within protected and surrounding waters of the tropics and temperate regions for both climate change scenarios (see Table S3 for full statistical analyses).
For the tropics, where most marine reserves are present (total number of grid cells 12,024), we found a significant difference (p < 0.05) in the distribution of biomass change direction for all combinations of grid cell type, time frame, and climate change scenario. Overall, it is expected that both protected and surrounding waters of the tropics lose biomass regardless of the climate change scenario (Fig. 3). Similarly, southern (total number of grid cells 4,332) and northern (total number of grid cells 1,036) temperate biomes presented significant (p < 0.05) differences in the distribution of the biomass change direction for all but one case. The only case where no significant difference is found (χ2 = 1.130841, df = 1, p = 0.29) is in northern temperate protected areas at the end of the 21st century under a low emission climate change scenario. While southern protected and surrounding waters tend to increase in biomass, northern waters are projected to decrease in biomass, regardless of the climate change scenario (Fig. 3). It is worth noting the lower number of marine reserves (and thus, grid cells) in the temperate north relative to other biomes. Finally, results in the southern polar biome are mixed. Here, most protected grid cells do not present a significant difference (p > 0.05) in the distribution of biomass change direction, except for the end of the 21st century under a high emission climate change scenario (χ2 = 17.6177606, df = 1, p = 0.3). However, all surrounding cells show a significant difference in the distribution of biomass change direction (p > 0.05), regardless of the time frame or climate change scenario. This result could be influenced by the ecological complexity of the biome as well as the characteristics of the few species modeled for the region and their associated fisheries (e.g., krill).
Fig. 3.
Finally, to evaluate if a world threatened by climate change will be better off with marine reserves in terms of fish biomass than a world without them (question iii), we compared the projected biomass with a version of the DBEM that did not incorporate marine reserves (Fig. 1.7). While comparing absolute biomass would be misleading (see the Methods section), we compare the biomass difference between conservation scenarios for each time frame and assess whether the difference expands or contracts. Under a low emission scenario, the difference in biomass between the conservation scenarios at present time is 30% (i.e., on average, there was 30% more biomass in the conservation scenario due to the presence of marine reserves). By 2050, that difference increased to 31% and by the end of the century to 34% suggesting that marine reserves are positively impacting biomass. However, under RCP 8.5 the difference is 30% for all time frames, suggesting that marine reserves alone cannot ameliorate climate change impacts under a high emission scenario.
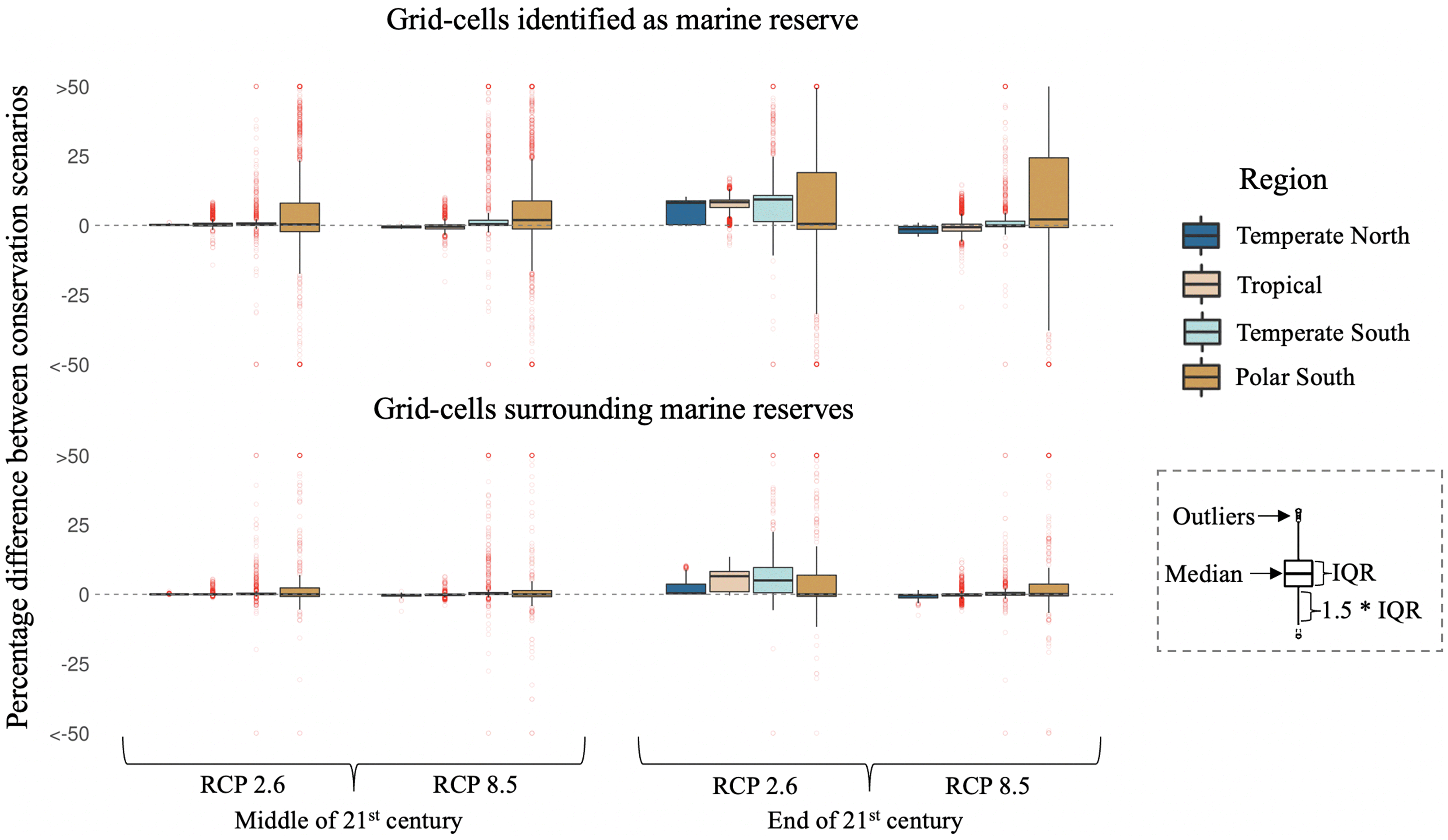
We also compared the differences between conservation scenarios in terms of percentage change in biomass relative to today. By the mid-21st century, protected cells from the conservation scenario average between 2% (s.d. 18%) under a low emission scenario and 3% (20%) under a high emission scenario more biomass change than the non-conservation scenario. Toward the end of the 21st century, the difference increases to 9% (s.d. 20%) under low emissions and decreases to 5% (s.d. 25%) under high emissions. While similar, albeit smaller, trends are found for surrounding grid cells, no difference in biomass change between conservation scenarios is found for unprotected grid cells, for all but the end of the 21st century under a low emission climate change scenario (Table S4). The southern temperate and polar protected and surrounding waters present the largest average gains for the conservation scenario in terms of biomass, especially toward the end of the century (Fig. 4). In contrast, tropical protected and surrounding waters show no apparent difference in (average) biomass change between conservation scenarios for all but the low emission scenario where they outperform the non-conservation scenario toward the end of the 21st century (Fig. 4). Although the conservation scenario outperformed the non-conservation scenario on a global aggregate, some grid cells (n = 4295; 2.5% of all grids) had a negative percentage difference between conservation scenarios (Fig. 4). In all of these cases, there was a higher starting biomass in the conservation scenario, leading to larger differences in percentage change in biomass compared to the non-conservation scenario that had relatively less initial biomass to lose (Table S5). Yet, in all cases, absolute biomass was still larger under the conservation scenario by both the mid and the end of the 21st century.
Fig. 4.
Discussion
We sought to understand how the biomass of marine reserves and its surrounding waters will be impacted by climate change globally and across different ecological regions. Our simulations suggest that climate change will have an overall negative impact on fish biomass within marine reserves and their surrounding waters throughout the 21st century. However, the impact varies from large losses in fish biomass in some tropical reserves to fish biomass gains in higher latitudes (Fig. 2). This pattern is especially pronounced under a high emission scenario (RCP 8.5) suggesting that some reserves (mostly tropical) will not be able to fully protect shifting species under extreme climate change. Yet, while tropical marine reserves may suffer the most under climate change, they could still provide benefits to communities in the future (McLeod et al. 2009; Davies et al. 2017). Overall, well-managed marine reserves are expected to continue to provide other benefits under climate change such as refugia area from the effects of fishing leading to older and larger (hence, more resilient) populations, the protection of ecosystem’s resilience and diversity, and carbon sequestration, thus promoting adaptation of socioecological systems to climate change (Micheli et al. 2012; Roberts et al. 2017; Jacquemont et al. 2022).
Climate change impacts to marine reserves will differ across ecological regions. Tropical marine reserves might not fully combat the expected range shifts of marine species toward more suitable habitats. Addressing such deficiency is critical as species richness in the tropics is higher (Reygondeau 2019), marine reserves are most abundant (UNEP-WCMC 2019), and climate change is expected to have some of the largest impacts on marine ecosystems in this region (IPCC 2019). While at localized levels tropical marine reserves could still provide some benefits, especially when marine species follow local temperature gradients away from the poles (Clarke et al. 2021), emissions mitigation will be required to enhance the protection of tropical biodiversity. As climate change pushes species away from the tropics, temperate marine reserves may be even more important as corridors for protecting biodiversity (Sullivan-Stack et al. 2022). Thus, the low number of marine reserves in the global northern temperate biome (UNEP-WCMC 2019) may need to be revised to protect biodiversity in this region. The presence of marine reserves in the polar regions may have positive outcomes for established species and future biomass moving into the region (Dahood et al. 2020) while counteracting an increase in fishing pressure (Watters et al. 2020). It is important to keep in mind the high uncertainty of climate change projections, and thus coupled ecological models, especially for polar regions (Bryndum-Buchholz et al. 2019; IPCC 2019). Accounting for such uncertainties while creating alternative designs of marine reserves that are climate resilient will help protect biomass from the impacts of climate change globally (Bryndum-Buchholz et al. 2022). Such designs include dynamic marine reserves that shift their borders following the redistribution of species under climate change (Maxwell et al. 2020; Cashion et al. 2020), large reserves (Davies et al. 2017), and networks of protected areas (McLeod et al. 2009; Sala et al. 2021).
We also sought to understand whether a changing world with marine reserves would be better off in terms of fish biomass than a world without the protection of the marine reserves. We found that a world with marine reserves will still outperform a world without them. However, the benefits of marine reserves are reduced under a high emission scenario. Under a high emission scenario, marine reserves will not be able to fully combat the major losses to biomass as a result of extreme climate change. Regardless of whether it is plausible or not to reach the warming level set by the high emission RCP 8.5 scenario (Burgess et al. 2022; Pielke et al. 2022), any greenhouse gas mitigation is expected to lead to less extreme changes in biomass, especially after the mid-21st century (Frölicher et al. 2016). Current global agreements, such as the Paris Agreement and UN Conference of Parties, are ever more important to not only keep global temperature increases below 2 °C but also promote the effectiveness of current marine reserves and achieve global sustainable development targets. Regionally speaking, having marine reserves is increasingly more beneficial in higher latitudes as species move from the tropics to polar regions and decrease in the tropics in an extreme (high emission) scenario (Cheung et al. 2009b; Cheung, Reygondeau, et al. 2016). The projected regional differences in biomass changes in protected waters can be attributed to the parallel change of several ocean variables (e.g., temperature, dissolved oxygen) from historical variability (Bruno et al. 2018). In theory, a world with marine reserves also means that surrounding waters will benefit from more biomass through spillover (Halpern et al. 2009). Our model supports this theory resulting in less losses in biomass in waters directly surrounding marine reserves in comparison to when no marine reserve is implemented, under future climate change scenarios. However, the spillover effect continues to be a source of contention as recent studies have noted that protected areas provide benefits to biomass within their borders, but that their contributions to local and regional population-wide effects are less evident (Ovando et al. 2021).
Marine reserves may ultimately support the desired goals of healthier and more diverse ecosystems under climate change; however, socioecological contexts must be considered when designing, maintaining, and implementing them (Charles and Wilson 2009; Dalton et al. 2015; Grorud-Colvert et al. 2021; Jacquemont et al. 2022). Reserves can be used to conserve core habitat for endangered species and may be used for recreation that supports marine conservation (Marcos et al. 2021). Reserves can have multiple goals beyond biomass or biodiversity conservation and climate change mitigation (Dalton et al. 2015), and these differential goals should be considered when designing and examining their effectiveness. Nonetheless, reserves may not always be the best approach (or only) to follow (Hilborn et al. 2004). For example, the implementation of a marine reserve on previously fished waters will likely displace fishing effort to surrounding regions (rather than eliminating it). This effect will often lead to overfishing in surrounding waters, potentially impacting the efficiency of the reserve and surrounding fisheries (Gilman et al. 2019; Cashion et al. 2020; Hilborn and Sinclair 2021). Thus, the implementation of marine reserves needs to be under a holistic ocean governance framework. Our hypothetical world requires sustainable fishing across the globe and that marine reserves are well enforced to isolate the effects of climate change. While these sound like utopian assumptions, the cost for supporting a network of marine reserves has been estimated to be less than that spent globally on harmful subsidies to industrial fisheries (Balmford et al. 2004), which also directly threaten ocean sustainability (Sumaila et al. 2021). Nevertheless, such assumptions mean that our model does not account for illegal fishing, enforcing of rules, nor potential displacement of fishing effort by marine reserves. Protecting ocean waters as a global responsive conservation strategy, especially in light of climate change and mass extinction, is an important contribution to the goals of the CBD which seeks to increase global protection coverage to 30% (CBD 2021) as well as achieving target 14 – Life Below Water of the Sustainable Development Goals (United Nations 2018). For marine conservation to thrive, ocean governance needs to be improved globally, while also considering the complexity of local socio-ecological contexts (Edgar et al. 2014; Ban et al. 2018; Sullivan-Stack et al. 2022; Hilborn and Sinclair 2021). The current conservation agenda should consider not only this but also the impacts of climate change to promote climate resilient marine reserves if we are to maintain and recover biodiversity in the ocean (IPBES 2019) while not negatively impacting other socioecological systems.
Conclusion
Our study shows the potential impacts of climate change to global marine reserves. Our findings emphasize the need to design climate resilient marine reserves to maintain and recover biodiversity in the ocean. Our results are the product of a hypothetical case study where marine reserves are well managed, enforced, and their benefits extend beyond their borders. In practice, reserves have complex socioecological dynamics with more nuances. The replication of our study at local levels where finer data and more knowledge of the socioecological system are available can better inform policy and management decisions. Such studies should focus on incorporating fleet dynamics, illegal fishing, and account for spatially explicit no-take proportion of the protected area to better quantify the benefits of marine reserves under climate change. Thoughtful future designations of marine reserves are needed not only in areas where the effects of climate change may be severe but also in areas where biomass will most likely be moving in the future, while accounting for the uncertainty of a changing world.
Acknowledgments
We thank Gabriel Reygondeau and Kelsey Roberts for helpful comments and discussions on the manuscript. We are also thankful to the anonymous reviewers whose comments substantially improved this manuscript. This work was supported by the National Socio-Environmental Synthesis Center (SESYNC) under funding received from the National Science Foundation DBI-1639145.
Compelling interests
The authors declare there are no competing interests.
References
Balmford A, Gravestock, P, Hockley, N, McClean, CJ, and Roberts CM. 2004 The worldwide costs of marine protected areas. Proceedings of the National Academy of Sciences, 101(26): 9694–9697.
Ban NC, Frid A, Reid M, Edgar B, Shaw D, and Siwallace P. 2018. Incorporate Indigenous perspectives for impactful research and effective management. Nature Ecology & Evolution, 2(11): 1680–1683.
Burgess MG, Becker SL, Langendorf RE, Fredston-Hermann A, and Brooks CM. 2022. Perspectives on most plausible climate futures, and recommendations for using scenarios in fisheries and aquatic conservation research. SorcArXiv [Preprint] 31 pages.
Bruno JF, Bates AE, Cacciapaglia C, Pike EP, Amstrup SC, van Hooidonk R, et al. 2018. Climate change threatens the world’s marine protected areas. Nature Climate Change, 8(6): 499–503.
Bryndum-Buchholz A, Boerder K, Stanley RRE, Hurley I, Boyce DG, Dunmall, KM. et al. 2022. A climate-resilient marine conservation network for Canada. FACETS, 7(1): 571–590.
Bryndum-Buchholz A, Tittensor DP, Blanchard JL, Cheung WWL, Coll M, Galbraith ED. et al. 2019. Twenty-first-century climate change impacts on marine animal biomass and ecosystem structure across ocean basins. Global Change Biology, 25(2): 459–472.
Cashion T, Nguyen T, ten Brink T, Mook A, Palacios-Abrantes J, and Roberts SM. 2020. Shifting seas, shifting boundaries: Dynamic marine protected area designs for a changing climate. PLoS ONE 15(11): e0241771.
CBD. 2021. First draft of the post-2020 global biodiversity framework. [online]: Available from cbd.int/doc/c/abb5/591f/2e46096d3f0330b08ce87a45/wg2020-03-03-en.pdf.
Charles A, and Wilson L. 2009. Human dimensions of Marine Protected Areas. ICES Journal of Marine Science, 66(1): 6–15.
Cheung WWL, Jones MC, Reygondeau G, Stock CA, Lam VWY, and Frölicher TL. 2016. Structural uncertainty in projecting global fisheries catches under climate change. Ecological Modelling, 325: 57–66.
Cheung WWL, Lam VWY, Sarmiento JL, Kearney K, Watson R, and Pauly D. 2009a. Projecting global marine biodiversity impacts under climate change scenarios. Fish and Fisheries, 10(3): 235–251.
Cheung WWL, Lam VWY, Sarmiento JL, Kearney K, Watson R, Zeller D et al. 2009b. Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Global Change Biology, 16(1): 24–35.
Cheung WWL, Reygondeau G, and Frölicher TL. 2016. Large benefits to marine fisheries of meeting the 1.5 °C global warming target. Science, 354(6319): 1591–1594.
Chirico AAD, McClanahan TR, and Eklöf JS. 2017. Community- and government-managed marine protected areas increase fish size, biomass and potential value. PLoS ONE 12(8): e0182342.
Clarke TM, Reygondeau G, Wabnitz C, Robertson R, Ixquiac-Cabrera M, López M, et al. 2021. Climate change impacts on living marine resources in the Eastern Tropical Pacific. Diversity and Distributions, 27(1): 65–81.
Costello MJ, and Ballantine B. 2015. Biodiversity conservation should focus on no-take Marine Reserves. Trends in Ecology and Evolution, 30(9): 507–509.
Crain CM, Halpern BS, Beck MW, and Kappel CV. 2009. Understanding and Managing Human Threats to the Coastal Marine Environment. Annals of the New York Academy of Sciences 1162(1): 39–62.
Dahood A, Mutsert K de, and Watters GM. 2020. Evaluating Antarctic marine protected area scenarios using a dynamic food web model. Biological Conservation, 251: 108766.
Dalton T, Forrester G, and Pollnac R. 2015. Are Caribbean MPAs making progress toward their goals and objectives? Marine Policy, 54: 69–76.
Davies TE, Maxwell SM, Kaschner K, Garilao C, and Ban NC. 2017. Large marine protected areas represent biodiversity now and under climate change. Scientific Reports, 7(1): 9569.
Dureuil M, Boerder K, Burnett KA, Froese R, and Worm B. 2018. Elevated trawling inside protected areas undermines conservation outcomes in a global fishing hot spot. Science, 362(6421): 1403–1407.
Edgar GJ, Stuart-Smith RD, Willis TJ, Kininmonth S, Baker SC, Banks S, et al. 2014. Global conservation outcomes depend on marine protected areas with five key features. Nature, 506(7487): 216–220.
Fredston-Hermann A, Gaines SD, and Halpern BS. 2018. Biogeographic constraints to marine conservation in a changing climate. Annals of the New York Academy of Sciences 1429(1): 5–17.
Friedlander AM, Golbuu Y, Ballesteros E, Caselle JE, Gouezo M, Olsudong D et al. 2017. Size, age, and habitat determine effectiveness of Palau’s Marine Protected Areas. PLoS ONE 12(3): e0174787.
Frölicher TL, Rodgers KB, Stock CA, and Cheung WWL. 2016. Sources of uncertainties in 21st century projections of potential ocean ecosystem stressors. Global Biogeochemical Cycles 30(8): 1224–1243.
Gattuso J-P, Magnan A, Billé R, Cheung WWL, Howes EL, Joos F, et al. 2015. Contrasting futures for ocean and society from different anthropogenic CO 2 emissions scenarios. Science, 349(6243): aac4722.
Grorud-Colvert K, Sullivan-Stack J, Roberts C, Constant V, Costa BH, Pike EP. et al. 2021. The MPA Guide: A framework to achieve global goals for the ocean. Science, 373(6560): eabf0861.
Gilman E, Kaiser MJ, and Chaloupka M. 2019. Do static and dynamic marine protected areas that restrict pelagic fishing achieve ecological objectives? Ecosphere, 10(12).
Halpern BS. 2003. Do reserves work and does reserve size matter? Ecological Applications 13(Suppl): 117–137.
Halpern BS. 2014. Making marine protected areas work. Nature, 506(7487): 167–168.
Halpern BS, Lester SE, and Kellner JB. 2009. Spillover from marine reserves and the replenishment of fished stocks. Environmental Conservation, 36(4): 268–276.
Hilborn R, and Sinclair ARE. 2021. Biodiversity protection in the 21st century needs intact habitat and protection from overexploitation whether inside or outside parks. Conservation Letters 14(4).
Hilborn R, Stokes K, Maguire J-J, Smith T, Botsford LW, Mangel M, et al. 2004. When can marine reserves improve fisheries management? Ocean & Coastal Management, 47(3–4): 197–205.
IPBES. 2019. Report of the plenary of the intergovernmental science-policy platform on biodiversity and ecosystem services on the work of its seventh session. Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, 47.
IPCC. 2019. IPCC special report on the ocean and cryosphere in a changing climate. [online]: Available from ipcc.ch/srocc/.
IPCC. 2022. Climate Change 2022: Impacts, Adaptation and Vulnerability. Intergovernmental Panel on Climate Change, Working Group II, p. 3675. [online]: Available from ipcc.ch/report/ar6/wg2/downloads/report/IPCC_AR6_WGII_FinalDraft_FullReport.pdf.
Jacquemont J, Blasiak R, Cam CL, Gouellec ML, and Claudet J. 2022. Ocean conservation boosts climate change mitigation and adaptation. One Earth, 5(10): 1126–1138.
Komsta L, and Novomestky F. 2015. Package momens; Moments, cumulants, skewness, kurtosis and related tests R:GPL–2 | GPL–3.
Marcos C, Díaz D, Fietz K, Forcada A, Ford A, García-Charton JA, et al. 2021. Reviewing the ecosystem services, societal goods, and benefits of marine protected areas. Frontiers in Marine Science, 8: 613819.
Maxwell SM, Gjerde KM, Conners MG, and Crowder LB. 2020. Mobile protected areas for biodiversity on the high seas. Science, 367(6475): 252–254.
McLeod E, Salm R, Green A, and Almany J. 2009. Designing marine protected area networks to address the impacts of climate change. Frontiers in Ecology and the Environment, 7(7): 362–370.
Meinshausen M, Smith SJ, Calvin K, Daniel JS, Kainuma MLT, Lamarque J-F, et al. 2011. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Climatic Change, 109(1–2): 213–241.
Micheli F, Halpern BS, Botsford LW, and Warner RR. 2004. Trajectories and correlates of community change in no-take marine reserves. Ecological Applications, 14(6): 1709–1723.
Micheli F, Saenz-Arroyo A, Greenley A, Vazquez L, Espinoza Montes JA, Rossetto M et al. 2012. Evidence that marine reserves enhance resilience to climatic impacts. PLoS ONE, 7(7): e40832.
O’Connor MI, Bruno JF, Gaines SD, Halpern BS, Lester SE, Kinlan BP. et al. 2007. Temperature control of larval dispersal and the implications for marine ecology, evolution, and conservation. Proceedings of the National Academy of Sciences, 104(4): 1266–1271.
Ovando D, Caselle JE, Costello C, Deschenes O, Gaines SD, Hilborn R. et al. 2021. Assessing the population-level conservation effects of marine protected areas. Conservation Biology, cobi.13782.
Pielke Jr R, Burgess MG, and Ritchie J. 2022. Plausible 2005–2050 emissions scenarios project between 2 °C and 3 °C of warming by 2100. Environmental Research Letters, 17(2): 024027.
Pinheiro J, and Bates D. 2020. Package nlme; Linear and Nonlinear Mixed Effects Models R (> = 3.4.0):GPL (> = 2) [online]: Available from svn.r-project.org/R-packages/trunk/nlme/
Poloczanska ES, Burrows MT, Brown CJ, García Molinos J, Halpern BS, Hoegh-Guldberg O, et al. 2016. Responses of marine organisms to climate change across oceans. Frontiers in marine science 3. [online]: Available from journal.frontiersin.org/Article/10.3389/fmars.2016.00062/abstract
Reygondeau G. 2019. Current and future biogeography of exploited marine groups under climate change. Predicting Future Oceans. Elsevier. pp. 87–101. [online]: Available from linkinghub.elsevier.com/retrieve/pii/B9780128179451000095
Riahi K, Rao S, Krey V, Cho C, Chirkov V, Fischer G, et al. 2011. RCP 8.5—A scenario of comparatively high greenhouse gas emissions. Climatic Change, 109(1–2): 33–57.
Roberts CM, O’Leary BC, McCauley DJ, Cury PM, Duarte CM, Lubchenco J, et al. 2017. Marine reserves can mitigate and promote adaptation to climate change. Proceedings of the National Academy of Sciences, 114(24): 6167–6175.
Sala E, Mayorga J, Bradley D, Cabral RB, Atwood TB, Auber A, et al. 2021. Protecting the global ocean for biodiversity, food and climate. Nature, 592(7854): 397–402.
Sumaila UR, Skerritt DJ, Schuhbauer A, Villasante S, Cisneros-Montemayor AM, Sinan H, et al. 2021. WTO must ban harmful fisheries subsidies. Science, 374(6567): 544.
Sullivan-Stack J, Aburto-Oropeza O, Brooks CM, Cabral RB, Caselle JE, Chan F, et al. 2022. A Scientific Synthesis of Marine Protected Areas in the United States: Status and Recommendations. Frontiers in Marine Science, 9: 849927.
Schiff K, Luk B, Gregorio D, and Gruber S. 2011. Assessing water quality in Marine Protected Areas from Southern California, USA. Marine Pollution Bulletin, 62(12): 2780–2786.
Song AM, Scholtens J, Stephen J, Bavinck M, and Chuenpagdee R. 2017. Transboundary research in fisheries. Marine Policy 76: 8–18.
Tittensor DP, Novaglio C, Harrison CS, Heneghan RF, Barrier N, Bianchi D, et al. 2021. Next-generation ensemble projections reveal higher climate risks for marine ecosystems. Nature Climate Change, 11: 973–981.
UNEP-WCMC. 2019. User manual for the world database on protected areas and world database on other effective area- based conservation measures: 1. 6. UNEP-WCMC: Cambridge, UK. [online]: Available from wcmc.io/WDPA_Manual
United Nations. 2018. Sustainable development goals. 17 Goals to Transform our World. [online]: Available from un.org/sustainabledevelopment/sustainable-development-goals/
van Vuuren DP, Stehfest E, den Elzen MGJ, Kram T, van Vliet J, Deetman S, et al. 2011. RCP2.6: exploring the possibility to keep global mean temperature increase below 2 °C. Climatic Change, 109(1–2): 95–116.
Watters GM, Hinke JT, and Reiss CS. 2020. Long-term observations from Antarctica demonstrate that mismatched scales of fisheries management and predator-prey interaction lead to erroneous conclusions about precaution. Scientific Reports, 10(1): 2314.
Zeller D, Palomares MLD, Tavakolie A, Ang M, Belhabib D, Cheung WWL, et al. 2016. Still catching attention: Sea Around Us reconstructed global catch data, their spatial expression and public accessibility. Marine Policy, 70: 145–152.
Supplementary material
Supplementary Material 1
- Download
- 2.25 MB
Information & Authors
Information
Published In

FACETS
Volume 8 • Number 1 • January 2023
Pages: 1 - 16
Editor: S.J. Cooke
History
Received: 26 April 2022
Accepted: 14 November 2022
Version of record online: 7 April 2023
Copyright
© 2023 Palacios-Abrantes et al. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Key Words
Sections
Subjects
Plain Language Summary
Using computer modeling to simulate a future with and without marine reserves under climate change
Authors
Author Contributions
JP-A, SMR, TB, TC, AM, and TN conceived and designed the study.
All performed the experiments/collected the data.
JP-A, SMR, TB, TC, AM, and TN analyzed and interpreted the data.
All contributed resources.
All drafted or revised the manuscript.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Juliano Palacios-Abrantes, Sarah M. Roberts, Talya ten Brink, Tim Cashion, William W.L. Cheung, Anne Mook, and Tu Nguyen. 2023. Incorporating protected areas into global fish biomass projections under climate change. FACETS.
8(): 1-16. https://doi.org/10.1139/facets-2022-0101
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item