River influence on mercury bioaccumulation in the coastal food web of Eeyou Istchee, James Bay, Canada
Abstract
Spatial patterns of bioaccumulated mercury were evaluated in coastal marine food webs of east Hudson Bay and east James Bay in the boreal subarctic of Canada. Two marine species, blue mussels (Mytilus edulis) and common eider ducks (Somateria mollissima) that consume mussels, were collected by a regional community-based monitoring network established in five communities. Stable isotope tracers (carbon, nitrogen, sulfur, and mercury) were measured to evaluate environmental drivers of mercury spatial patterns. Mercury concentrations of blue mussels and common eiders were twofold and fivefold higher, respectively, on the James Bay coast near the community of Chisasibi compared to sites in east Hudson Bay. Liver and muscle mercury concentrations of eiders from James Bay are among the highest values reported for the circumpolar subarctic and Arctic. Multiple lines of evidence (mercury spatial patterns, crustal elements in blue mussels, and mercury isotope values of common eiders) suggest elevated mercury in the coastal food web of east James Bay may be due to mercury loading from the La Grande River, which drains one of the largest hydroelectric developments in the world. These findings highlight the importance of further research on environmental processes linking large rivers to mercury bioaccumulation in northern coastal food webs.
Graphical Abstract
Introduction
Hudson Bay, including its southern arm James Bay, is the world’s largest polar inland sea (Keller et al. 2014). It is home to Cree and Inuit, whose coastal communities rely on this vast ecosystem to harvest marine mammals, waterfowl, fish, and invertebrates (McDonald et al. 1997; NTK 2008; Hamilton 2011). The Hudson Bay and James Bay environments are being altered by human-driven stressors, both far and near, including climate warming, hydro-electric development, watershed alteration, and contaminant transport (Mallory et al. 2010; Stewart and Barber 2010; Hamilton 2011). Efforts are underway to develop inter-jurisdictional stewardship that addresses these complex transboundary issues with inclusion of community-based approaches and complementary Indigenous and scientific knowledge on cumulative impacts (NTK 2008; Hamilton 2011; Hudson Bay Consortium 2020). A community-driven research network has developed in association with Arctic Eider Society for east Hudson Bay and James Bay to measure and better understand large-scale cumulative environmental impacts, including changes in sea-ice (Arctic Eider Society 2024; Peck et al. 2022) and contaminants (Government of Canada 2022).
Among contaminants, mercury is a priority because this neurotoxin (more specifically the organic form methylmercury [MeHg]) biomagnifies through food webs and can reach elevated concentrations in top predator animals of Arctic marine ecosystems (Braune et al. 2015). In the marine environment, MeHg is produced by microbial methylation of inorganic mercury in sediments and the water column, and it is also formed via the degradation of dimethylmercury (Lehnherr 2014). Consumption of certain country foods, particularly marine mammals, can increase the risk of mercury exposure to humans (Lemire et al. 2015). Mercury enters Hudson Bay primarily via atmospheric deposition after long-range transport from distant emission sources as well as by river influx from vast watersheds (Hare et al. 2008). Hudson Bay receives inflow of fresh water (and mercury) from rivers that drain an area greater than one third of Canada (Déry et al. 2011), and such a large catchment drainage of mercury could potentially impact coastal food webs.
In northern Quebec, the La Grande hydroelectric complex is one of the largest hydro developments in the world, with river diversion and impoundments that power eight generating stations (HydroQuebec 2023). Infrastructure construction began in 1973 and expansions continued until as recently as 2012 (Bilodeau et al. 2017; HydroQuebec 2023). The flooding of labile organic matter in soils and vegetation when impoundments are created results in elevated microbial methylation of inorganic mercury to form MeHg (Porvari and Verta 1995; Hall et al. 2005). Monitoring of freshwater fish in older impoundments of the La Grande complex showed mercury contamination occurred after flooding with a return to background concentrations in fish muscle after two to three decades (Bilodeau et al. 2017). Coastal communities located in the region have raised concerns about the potential for mercury transport via ocean currents following discharge from the La Grande River to James Bay (Government of Canada 2022). The La Grande River contributes the largest river flux of MeHg to the east coast of James Bay, reflecting in part the high discharge from this system (Fink‐Mercier et al. 2022). The La Grande hydroelectric complex has altered the hydrology of the La Grande River by increasing freshwater discharge to the east coast of James Bay at the community of Chisasibi, which has affected sea grass beds (Murphy et al. 2021) and winter ice conditions (Peck et al. 2022). Winter discharge is tenfold higher (up to ∼5000 m3/s) compared with pre-development conditions (Déry et al. 2016; Peck et al. 2022).
Investigations of mercury bioaccumulation in marine biota of Hudson Bay have focused largely on temporal trends or broad-scale spatial patterns across the Arctic Ocean for seabirds, ringed seal (Pusa hispida), and polar bear (Ursus maritumus) (e.g., Braune et al. 2014; Brown et al. 2016; Mallory et al. 2017; Morris et al. 2022). This research has shown mercury concentrations tend to be lower in tissues of Hudson Bay polar bear and ringed seal relative to higher latitude locations in the Arctic Archipelago (St. Louis et al. 2011; Brown et al. 2016). Long-term seabird monitoring in northern Hudson Bay (at Coats Island) indicates that inter-annual variation in mercury concentrations of eggs is related to shifts in parental diet and climate (Braune et al. 2014; Morris et al. 2022). Limited information is available on spatial variation of mercury bioaccumulation at finer geographic scales within Hudson Bay and on local environmental drivers of MeHg uptake at the base of the marine food web.
Here, we report on spatial patterns of mercury concentrations of marine biota of east Hudson Bay and James Bay. Characterization of mercury biomagnification and trophic transfer through the east Hudson Bay food web is reported in a separate publication, which showed the importance of habitat-specific feeding and enhance mercury bioaccumulation in vertebrate species supported more by pelagic carbon (Rohonczy et al. 2024). In the present study, a suite of diet tracers (carbon, nitrogen, and sulfur stable isotope ratios) and source tracers (mercury stable isotopes and tissue burdens of other metals) were measured to evaluate environmental drivers of mercury spatial patterns. This study addresses a knowledge gap on environmental controls of mercury bioaccumulation in coastal marine ecosystems of the boreal subarctic in Canada.
Materials and methods
Study area and design
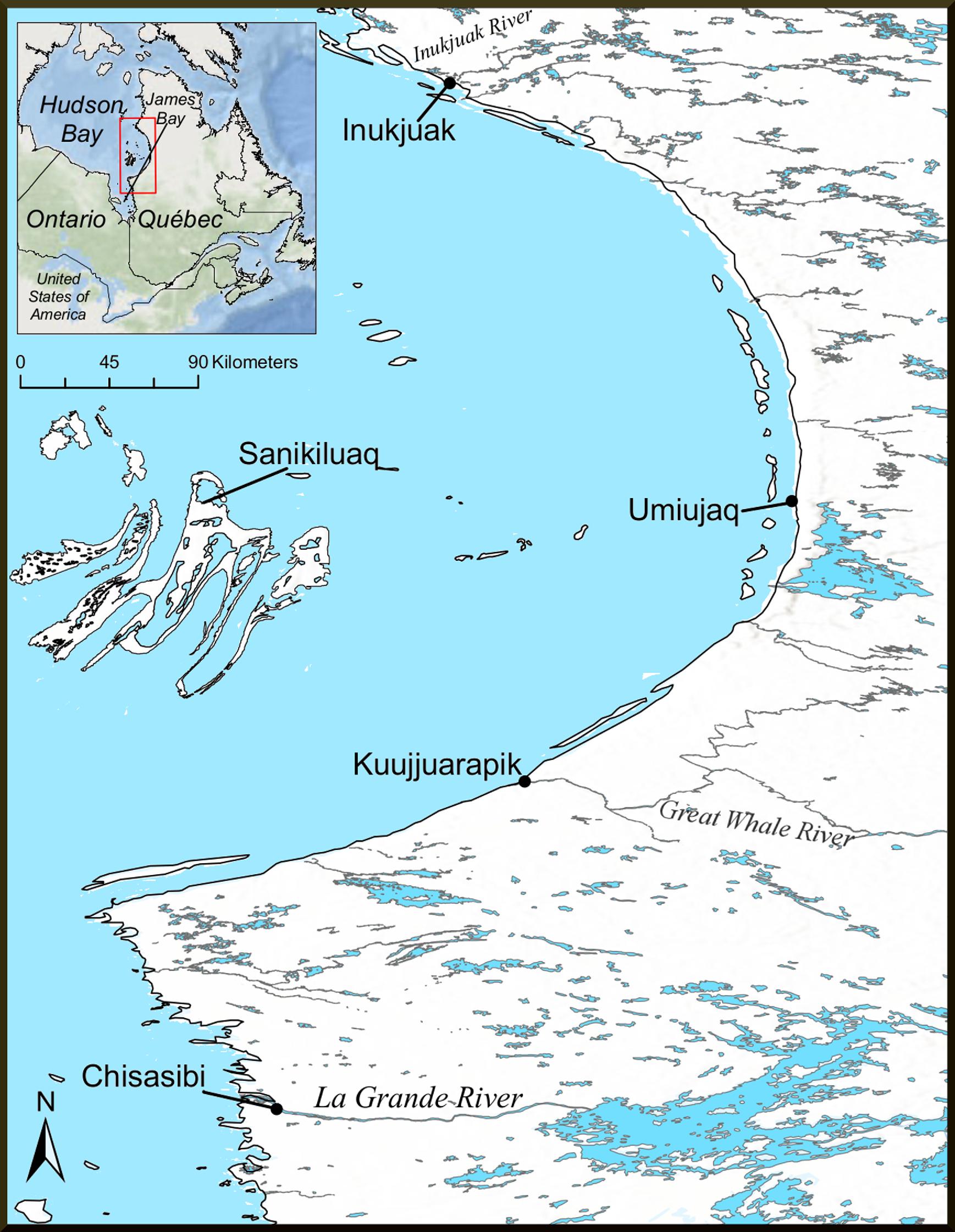
A regional community-driven research network (Arctic Eider Society 2024) participated in a study of metal concentrations in coastal food webs of four communities on east Hudson Bay (Sanikiluaq, Kuujjuarapik, Umiujaq, and Inukjuak) and the community of Chisasibi on east James Bay (Fig. 1). The southern portion of the study area is within Eeyou Istchee, the traditional territory of the Cree of Quebec, while the northern portion comprises Inuit homelands of Nunavik and Nunavut. Hudson Bay and James Bay are polar marine ecosystems with winter ice cover and ice-adapted marine animals typical of more northern latitudes including polar bear, Arctic cod (Boreogadus saida), and ringed seal (Stewart and Barber 2010). The region is also important for migratory birds, which use coastal habitats during the spring and summer seasons. More details about the environmental characteristics of the Hudson Bay complex can be found in Stewart and Lockhart (2005), Stewart and Barber (2010), and Keller et al. (2014).
Fig. 1.
Spatial patterns of mercury bioaccumulation were investigated using two bioindicator species found throughout the study area: blue mussel (Mytilus edulis) and common eider (Somateria mollissima). Blue mussel is a sedentary filter-feeding invertebrate that feeds predominately on pelagic rather than sediment carbon (Rosa et al. 2018), and it is widely studied around the world to monitor pollution (Beyer et al. 2017). Common eider is a diving sea duck that feeds on molluscs (including blue mussel), crustaceans, and echinoderms in benthic habitats (Ouellet et al. 2013). Common eiders in east Hudson Bay and east James Bay are the sedentaria subspecies, which are non-migratory and remain year-round by overwintering in open water of polynyas and leads of ice particularly around the Belcher Islands (Mallory et al. 2004; Goudie et al. 2020). In spring, the eiders disperse to breeding grounds on islands and in coastal areas of James Bay and Hudson Bay, where they remain for much of the open-water season to breed, rear their young, and molt (Goudie et al. 2020).
Blue mussels and common eiders were collected by local hunters near each of their communities. The collections were completed during the open water season (June to early December) between 2014 and 2017, though additional common eiders were also collected at Chisasibi in 2021 to increase the sample size. Blue mussels were collected by hand or trawl net from several near-shore sites per community. Ten individual mussels were collected per site to form a composite sample, and the number of composite blue mussel samples varied from 6 to 20 per community, representing sampling of multiple sites over a 2- or 3-year period. A total of 59 composite blue mussel samples were obtained for the study. Blue mussels were frozen whole in polyethylene zip bags and shipped to the National Wildlife Research Centre (NWRC, Ottawa, Canada) for laboratory processing. Common eiders were captured with a firearm from sites near each community, with sample sizes ranging from 14 to 24 birds over a 2- or 3-year period, except for Inukjuak where only 4 birds were collected in 1 year. Breast muscle and the liver were removed from each bird after collection, frozen in polyethylene zip bags, and shipped to NWRC for laboratory processing. A total of 77 common eiders were obtained for the study. The repeat sampling of blue mussel and common eider at each community incorporated both local spatial variation and inter-annual variation.
Laboratory methods
In the laboratory, biotic samples were homogenized and freeze-dried prior to chemical analysis. Blue mussels were de-shelled, and the whole-body soft tissue of 10 individuals from a site was homogenized with an Omni Mixer with wide window shafts to form a composite sample. Individual common eider livers were also mechanically homogenized while eider muscle samples were manually homogenized in their containers after drying. All samples were freeze-dried for a minimum of 48 h.
Total mercury (THg) was measured in blue mussel, eider liver, and eider muscle with a Direct Mercury Analyzer (Milestone Inc., Shelton, Connecticut, USA) at NWRC. For a small subset of samples (liver and muscle of 14 eiders collected in 2021), THg was measured following wet digestion by atomic absorption spectrometry at RPC laboratories (Fredericton, New Brunswick, Canada) due to lab closure at NWRC. Analytical duplicates were measured every 10 samples with a relative percent difference (RPD) of <10% (n = 14). During the project, six types of certified reference materials (CRMs) were analyzed showing mean recoveries of 86%–102% (n = 108; see Rohonczy et al. 2024, for details). An inter-laboratory comparison of eider mercury results from RPC with those from NWRC showed high precision (mean RPD = 10%, n = 10).
Methylmercury was also measured on a subset of blue mussel samples (n = 44) with a cold-vapour atomic fluorescence spectrometer at NWRC or Flett Research Ltd (Winnipeg, Manitoba, Canada). Samples were digested prior to analytical detection with nitric acid at NWRC and with KOH-methanol at Flett Research Ltd. Duplicate analytical measurements had a mean RPD = 4% (n = 7). Recoveries of CRMs averaged 90% (range of 85%–97%) for both DORM-3 fish protein (n = 4; National Research Council of Canada) and NIST 2976 mussel tissue (n = 6; National Institute of Science and Technology).
A suite of 24 elements was measured in blue mussel samples (n = 59) with an inductively coupled plasma mass spectrometer (ICP-MS) at NWRC or the Alberta Institute of Technology Futures (Vegreville, Alberta, Canada). Only elements that were above analytical detection (Ag, As, Ba, Cd, Co, Cu, Fe, Li, Mn, Mo, Ni, Pb, Se, Sr, U, V, and Zn) are reported here. Analytical duplicates were measured every 10 samples with deviations generally <10%. Mean recoveries of individual elements ranged from 92% to 105% for measurement of 2–8 types of CRMs, with the exception of Pb, which had a lower mean recovery (77%). Details on the quality assurance and quality control of measurements for individual elements are provided in the supplementary materials (Table S1).
Blue mussels and common eider muscle were analyzed for nitrogen, carbon, and sulfur isotope ratios on a DeltaPlus XP isotope ratio mass spectrometer interfaced to a Vario El III elemental analyzer via a Conflo II at the Ján Viezer Stable Isotope Laboratory at the University of Ottawa (Ottawa, Ontario, Canada). Isotope ratios are reported in delta notation as the per mil (‰) deviation from N2 (air) for nitrogen, Vienna Pee Dee Belemnite for carbon, and Vienna Canyon Diablo Troilite for sulfur. Analytical precision is typical <0.2‰ for carbon and nitrogen stable isotopes and <0.3‰ sulfur stable isotopes.
Mercury stable isotope ratios were measured on a subset of common eider muscle samples (n = 25) with a Nu Plasma II multi collector-ICP-MS instrument in the Water Quality Centre at Trent University (Peterborough, Ontario, Canada). Samples were digested using 3 mL of aqua regia in a 40 mL amber glass vessel by heating on a hot plate at 120 °C for 4 h. An ESI hydride ICP hydride generation system was used to generate a Hg0 vapor by the quantitative reduction of Hg(II) in solution using stannous chloride (3% w/v in 1 M HCl). Mercury isotope ratios are reported as the per mil deviation from the xxxHg/198 Hg ratio of the NIST SRM 3133 mercury standard. Results for replicate measurements of the RM-8610 cinnabar standard are provided in the supplementary materials (Table S2).
Data analysis
The raw concentration data for THg, MeHg, and other elements as well as carbon, nitrogen, and sulfur isotope ratios are available online in an open-access database (Government of Canada 2022), and summarized data for common eiders and blue mussels from Hudson Bay are also reported in Rohonczy et al. (2024). Spatial patterns of mercury concentrations in biota were examined by comparing community means using one-way Analysis of variance (ANOVA) tests. Data were log-transformed, as required, and probability values of pair-wise comparisons were Holm-adjusted to correct for the experiment-wise error rate. Diet influences on mercury concentrations of common eider were examined by testing Pearson correlations with carbon, nitrogen, and sulfur stable isotope ratios. Blue mussels are sedentary bio-indicators of metal bioaccumulation near the base of the food web, and therefore, their element concentrations were examined for potential spatial patterns in metal exposure and links with mercury bioaccumulation. A principal component analysis (PCA) of blue mussel element concentrations was performed in R program (R Core Team 2021) using the built-in prcomp function and plotted with the ggbiplot package. Pearson correlations were tested for associations of blue mussel THg and MeHg concentrations with other tissue element concentrations. Mercury sources and exposure pathways to common eider were investigated by comparing mercury stable isotope ratios between communities with one-way ANOVAs.
Results and discussion
Spatial variation in mercury bioaccumulation
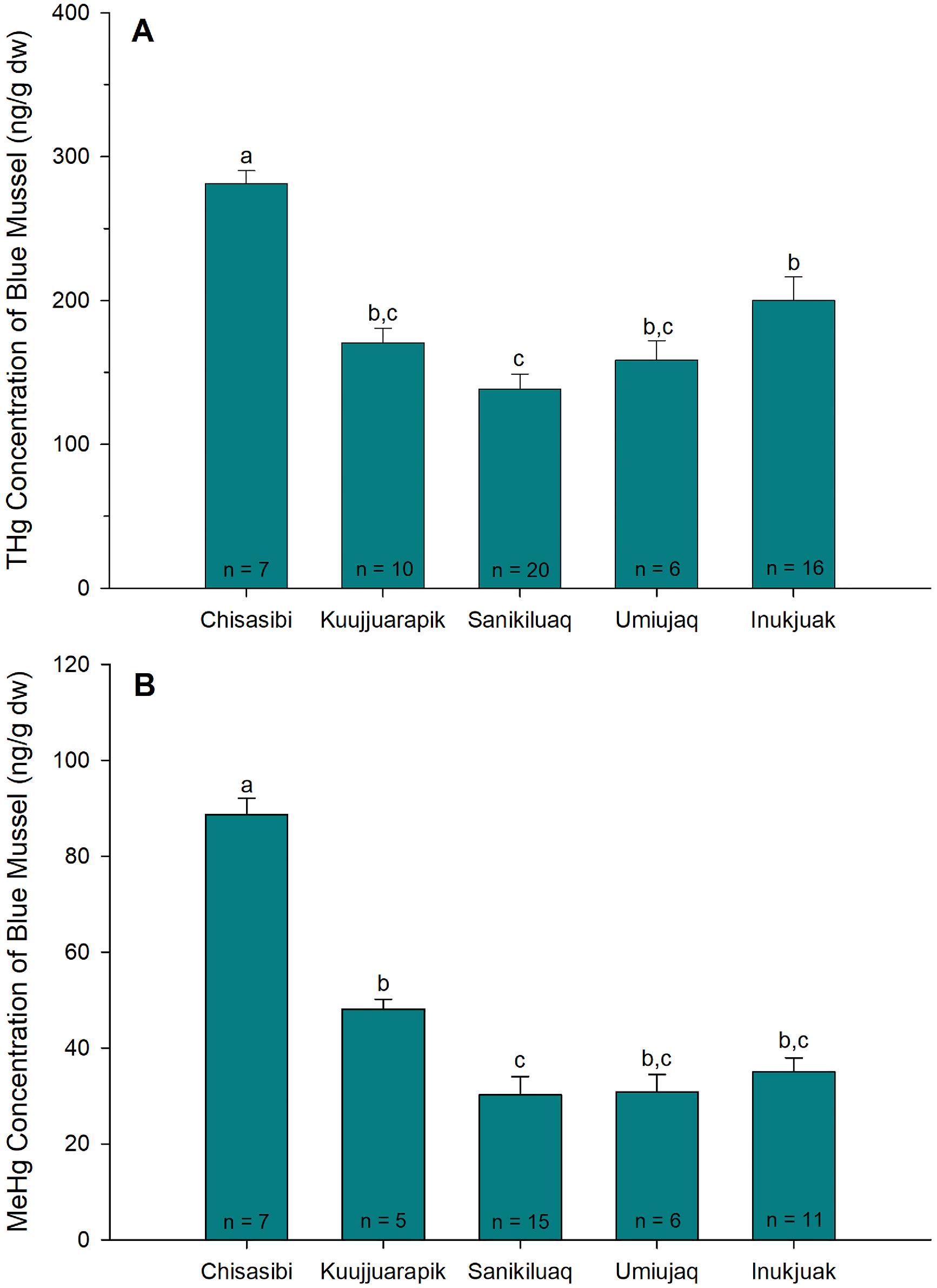
A clear spatial pattern was evident of higher mercury concentrations in blue mussels from near Chisasibi (Fig. 2). Those blue mussels had statistically higher THg and MeHg concentrations compared to mussels collected near four communities in east Hudson Bay (one-way ANOVAs, F4,58 = 12.6 (THg), F4,43 = 36.4 (MeHg), p < 0.001). There was approximately a twofold difference in mean THg and MeHg concentrations of blue mussels between Chisasibi and Sanikiluaq, the community where the lowest mercury concentrations were observed. In addition, the mean (± standard error (SE)) percent MeHg content of blue mussels was higher near Chisasibi (32% ± 1%) compared with three of the four other communities (Umiujaq, Sanikiluaq, and Inukjuak), where percent MeHg means ranged from 19% to 21% (Supplemental Fig. S1). The higher THg, MeHg, and percent MeHg of blue mussels near Chisasibi indicated greater mercury exposure near the base of the food web in that area of east James Bay.
Fig. 2.
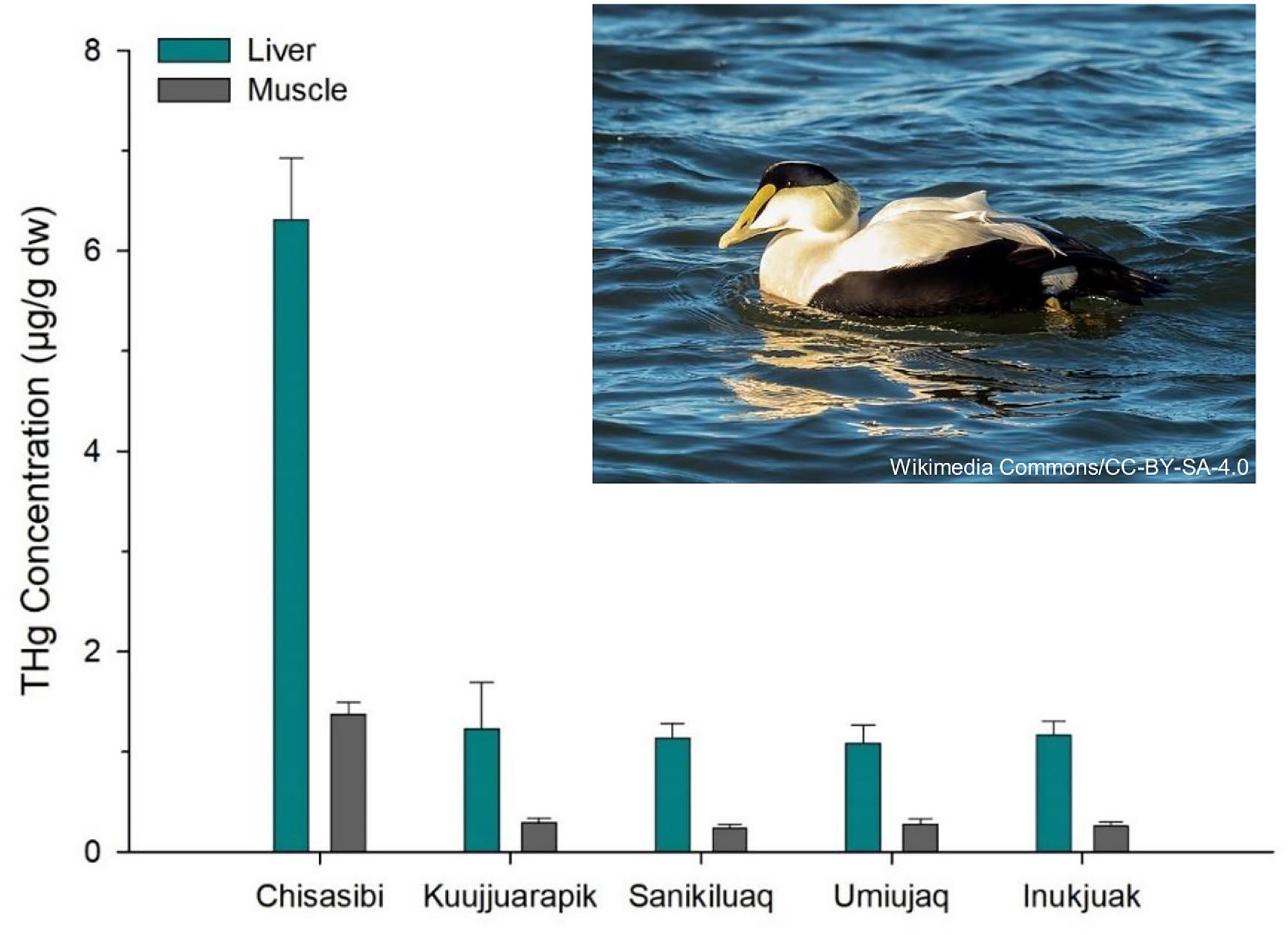
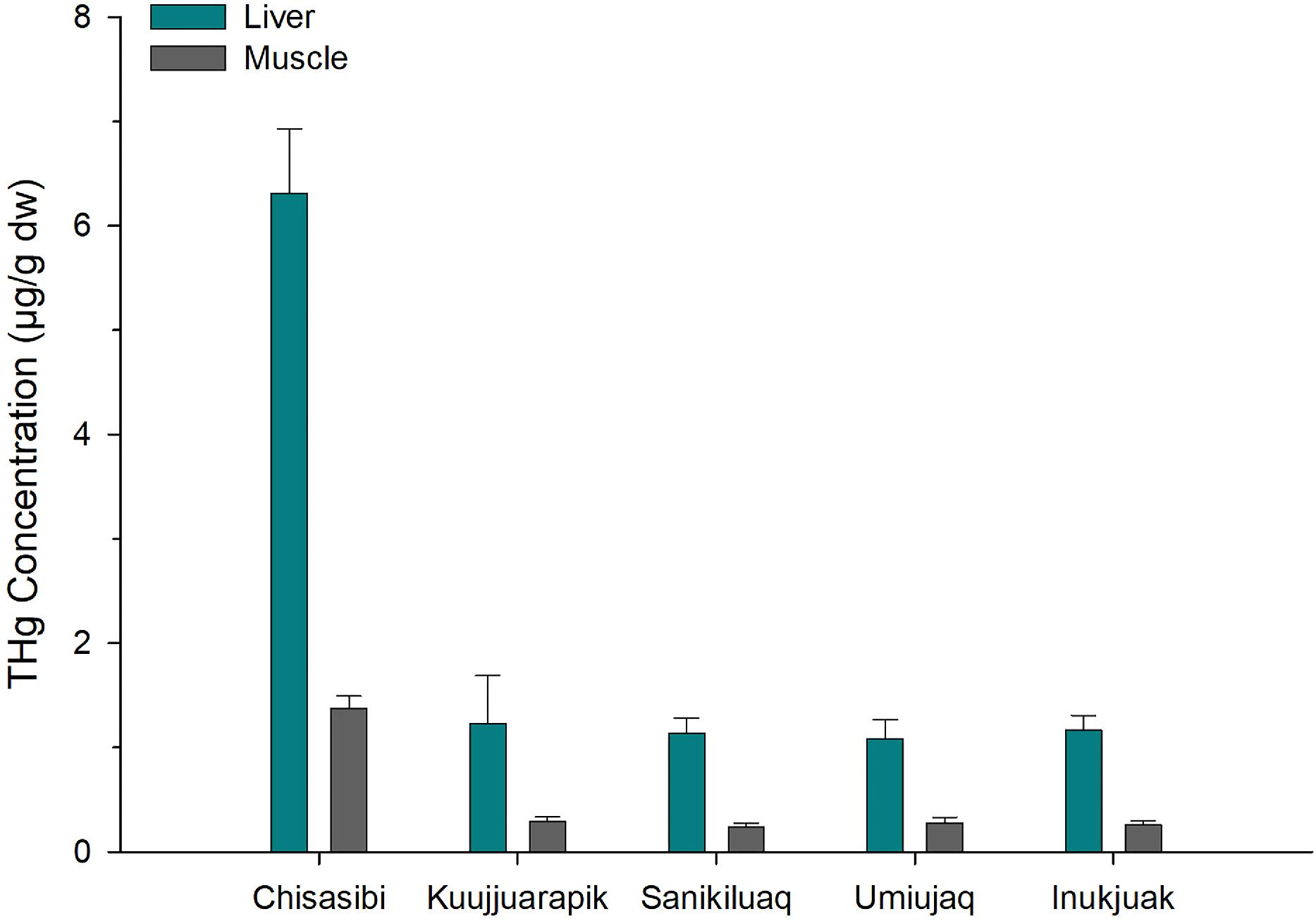
Common eiders, which feed on blue mussels, showed a corresponding spatial pattern in THg concentration (Fig. 3). Both liver and breast muscle of eider near Chisasibi had approximately fivefold higher THg compared to the four communities in east Hudson Bay (one-way ANOVAs, F4,76 = 23.4–33.2, p < 0.001). Concentrations of THg in eider were similar among the four Hudson Bay communities. For all birds, liver THg concentrations were higher than in muscle. Note that the THg of muscle is mainly in the form of MeHg while liver contains variable amounts of both inorganic mercury and MeHg (Chételat et al. 2020).
Fig. 3.
The higher THg concentrations of blue mussels near Chisasibi (mean ± SE = 0.28 ± 0.01 µg/g) were above median and mean values, though within the range, of published data reported in other geographic regions for this widely distributed species of marine bivalve. In the Estuary and Gulf of St. Lawrence in eastern Canada, THg concentrations of blue mussels ranged from 0.06 to 0.51 µg/g with a grand mean of 0.17 µg/g and a median of 0.16 µg/g (Cossa and Tabard 2020). At 20 coastal sites in the North Atlantic subarctic and Arctic, blue mussel THg concentrations were typically <0.1 µg/g and the maximum site mean was 0.23 µg/g (Jörundsdóttir et al. 2014). In the Baltic Sea, median THg concentrations of blue mussels were ≤0.08 µg/g though maximum values reached ∼0.3–0.5 µg/g (converted from wet weight values assuming 87% moisture content) (Dietz et al. 2021). Blue mussels on the coast of Massachusetts and Maine in the United States had mean THg concentrations 0.13 and 0.07 µg/g, respectively (Meattey et al. 2014). Blue mussels are widely used as biosentinels of coastal marine pollution around the globe and their mercury burdens reflect local environmental contamination, although other biological and environment factors such as growth and seasonal variability can also be influential (Beyer et al. 2017; Cossa and Tabard 2020).
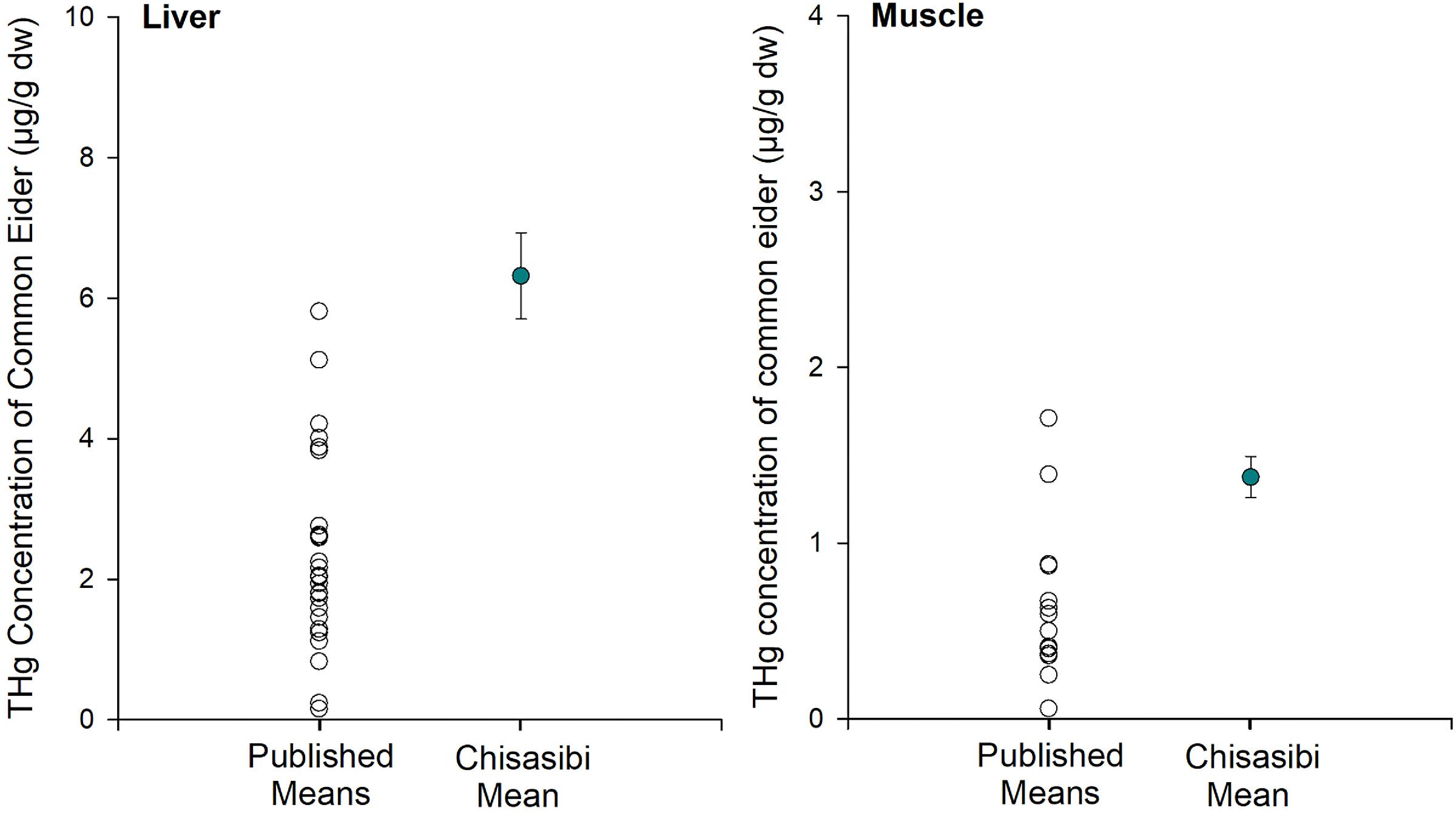
For common eider, the THg concentrations in liver and muscle from Chisasibi are among the highest reported for the circumpolar subarctic and Arctic (Fig. 4). Published THg concentrations were available from 23 sites for common eider liver and from 12 sites for muscle (data sources are provided in Supplemental Table S3). Much of these data were compiled from previous reviews and spatial studies of mercury concentrations in common eiders (Braune et al. 1999; Mallory et al. 2004; Mallory et al. 2017). Published site means ranged from 0.15 to 5.81 µg/g for liver and from 0.06 to 1.71 µg/g for muscle. In comparison to those values, the eiders collected near Chisasibi had the highest mean THg concentration for liver (mean ± SE = 6.31 ± 0.61 µg/g) and the second highest mean for muscle (mean ± SE = 1.37 ± 0.12 µg/g). Thus, the regionally elevated mercury concentrations of those common eiders are also exceptional more broadly in the circumpolar Arctic. While biological factors such as sex can influence mercury concentrations of common eider, differences between males and females are small relative to the large spatial variation observed in this study (Provencher et al. 2016). The eiders were collected near Chisasibi during their period of residency on James Bay, and their mercury burdens likely reflect local dietary exposure. The pattern is consistent with the elevated mercury in blue mussels (an important diet item for eider) from the same study area. The mercury concentrations of liver and muscle were compared to established toxicity risk thresholds for birds (calculated using blood equivalent concentrations, see Ackerman et al. 2016), which suggested no to low risk of mercury toxicity to the eiders (Chastel et al. 2022).
Fig. 4.
Dietary influences on mercury bioaccumulation of common eider
Carbon, nitrogen, and sulfur isotope ratios indicated an influence of diet on the THg concentrations of common eiders. Only minor differences were observed in mean δ13C values of eiders, ranging from −20.2 to −19.1‰ among the five communities (Supplemental Fig. S2). Despite the small range of δ13C values, carbon isotope ratios were negatively correlated with log-transformed THg concentrations of eider (muscle: Pearson r = −0.35, p = 0.002; liver: Pearson r = −0.47, p ≤ 0.001; n = 77). Little variation was observed in the mean δ15N values of eiders among communities (10.3–11.3‰), indicating a similar trophic position among birds collected in the study. There was no association between eider δ15N values and their log-transformed THg concentrations (Pearson correlation, p ≥ 0.10, n = 77). A larger range of sulfur isotope ratios was observed, with eider mean values ranging from 12.9 to 17.2‰ between communities and a similar range of variation (∼5–8‰) within most collection areas (Supplemental Fig. S2). The sulfur isotope ratios were not correlated with log-transformed THg concentrations of eider liver or muscle (Pearson correlation, p ≥ 0.14, n = 77). Thus, only the dietary carbon source of eiders was associated with THg concentrations in their tissues.
The carbon isotope values of common eiders were consistent with diving sea ducks feeding on a benthic diet in the study region (Rohonczy et al. 2024). Blue mussels, which primarily filter organic matter from the water column, had a mean δ13C (± SD) of −22.7 ± 0.6‰ (n = 59), while sea urchin, which consume organic matter at the ocean floor, had a mean δ13C of −18.6 ± 0.9‰ (n = 19) (as reported in Rohonczy et al. 2024). Considering those two end members, the negative correlation between eider THg concentrations and carbon isotope ratios suggests higher exposure to mercury occurred through greater consumption of marine invertebrates supported by water column organic matter, such as filter feeders. Other Arctic marine food web studies have similarly found that greater feeding of pelagic-derived carbon was associated with higher mercury concentrations in consumers (Cardona‐Marek et al. 2009; Hilgendag et al. 2022; Rohonczy et al. 2024). There was a notable absence of a freshwater organic matter signal in the stable isotope content of eider muscle. The δ13C values (>−21‰) did not suggest high consumption of terrestrial organic matter from river inputs, which has lower values close to −27‰ (Godin et al. 2017). Likewise, the δ34S values of eider muscle were >10‰, which are higher than values typically observed in freshwater biota (Chételat et al. 2020).
Environmental influences on food web uptake of mercury
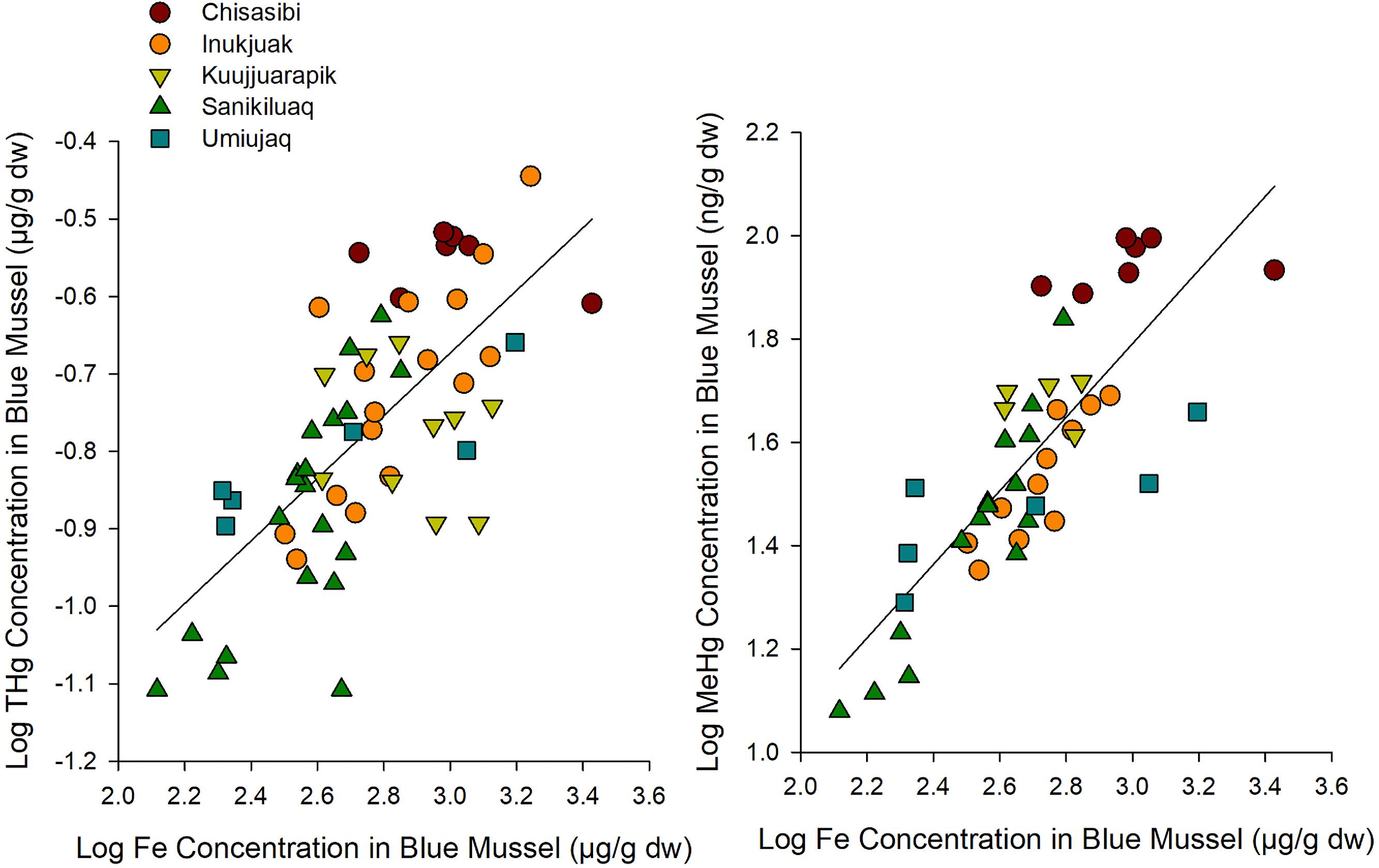
The elemental composition of blue mussels suggested environmental conditions varied among study sites. A PCA of tissue concentrations of 18 elements in blue mussels showed broadly similar burdens (as indicated by overlapping 95% confidence ellipses) for mussels near three Hudson Bay communities (Inukjuak, Umiujaq, and Sanikiluaq) and differences from mussels at Chisasibi and Kuujjuarapik (Supplemental Fig. S3). While many elements contributed to the PCA gradients in tissue concentrations, the THg of blue mussels was most strongly correlated with cobalt, iron, uranium, and vanadium (Pearson r = 0.67–0.71, p < 0.001, n = 59; Supplemental Fig. S4). Those elements were also strongly correlated with MeHg concentrations of blue mussels (Pearson r = 0.61–0.81, p < 0.001, n = 44). Iron was noteworthy in its strong association with THg and MeHg concentrations of blue mussels (Fig. 5). This co-variation of tissue element burdens provided an indication of possible environmental processes associated with greater mercury bioaccumulation in blue mussels. We hypothesize that the positive associations of mercury with iron and other crustal elements reflect the influence of river inputs on mussel exposure to metals.
Fig. 5.
Rivers transport vast amounts of materials to oceans, linking crustal weathering processes in terrestrial environments to ocean shelves (Andersen et al. 2007; Krachler and Krachler 2021). On a global scale, riverine sources of many elements (including cobalt, iron, vanadium, and uranium) are among the most important fluxes in ocean budgets (Andersen et al. 2007; Boyd and Ellwood 2010; Swanner et al. 2014; Schlesinger et al. 2017). River water contains organic matter and trace elements, which are distributed between dissolved, colloidal, and particulate phases. Estuaries are sinks for terrestrial organic matter and crustal elements transported by rivers because of particle settling and flocculation (Boyle et al. 1977; Demarty et al. 2021). The colloidal fraction of river water, rich in iron and carbon, is a dominant carrier of trace elements, including mercury (Jilbert et al. 2018; Demarty et al. 2021; Saniewska et al. 2022). When river water colloids interact with saline water in estuaries, flocculation occurs, causing a large fraction of trace metals and organic matter to be efficiently removed to sediments in the mixing zone (Jokinen et al. 2020; Khoo et al. 2022). Though few empirical measurements exist, high removal rates have been reported for riverine mercury entering estuaries due to flocculation and particle settling (Soerensen et al. 2016; Demarty et al. 2021; Saniewska et al. 2022). Model estimates suggest that 94% of mercury discharged from rivers is sequestered in estuary and coastal shelf sediments (Zhang et al. 2015). Together, the covariance of mercury with iron and other crustal elements in blue mussels across sites may be explained by exposure to river-transported elements that settle out on the coast. However, this hypothesis requires further study, and other factors could be influencing the patterns including differences in benthic substrates between Hudson Bay and James Bay, particularly the presence of tidal mud flats in James Bay.
Mercury sources and exposure pathways to common eider
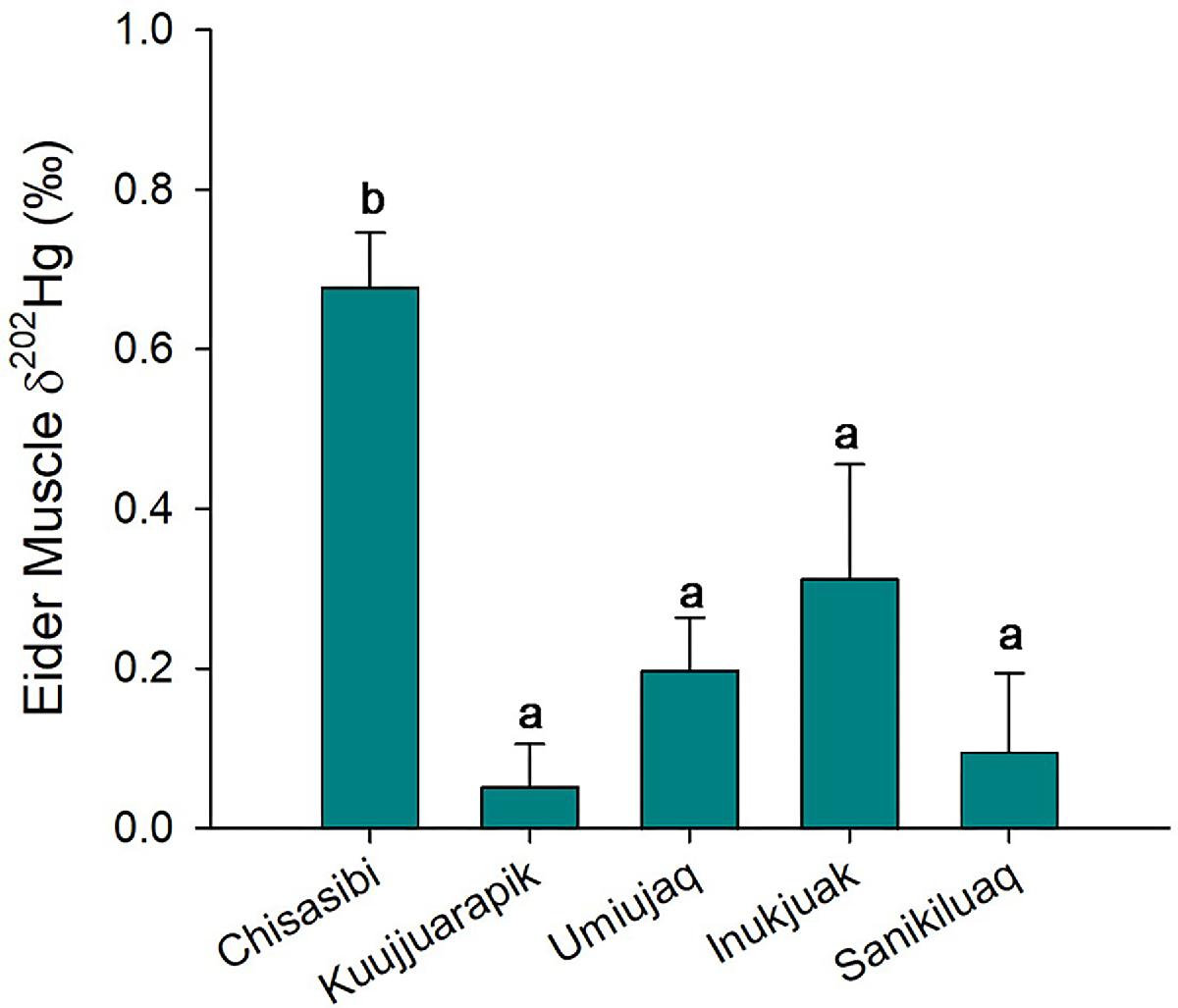
Mercury isotope values of eiders from Chisasibi were different from other communities in the study area. The δ202Hg values of a subset of eider muscle samples (n = 25) from the five communities ranged from −0.29 to 0.85‰, and the Δ199Hg values ranged from 0.42 to 2.22‰. On average, eiders from Chisasibi had significantly higher δ202Hg values than those from the other four communities (Fig. 6; one-way ANOVA, p < 0.001, n = 25, Holm’s p ≤ 0.044). Similarly, the Δ199Hg values of Chisasibi eiders were significantly higher than those from three of the other communities (one-way ANOVA, p < 0.001, n = 25, Holm’s p ≤ 0.014 for Sanikiluaq, Kuujjuarapik, and Inukjuak). This spatial variation of mercury isotope ratios indicated different origins of bioaccumulated mercury in eiders from Chisasibi.
Fig. 6.
Variation in Δ199Hg values is caused by mass-independent fractionation during photoreduction of inorganic mercury and photochemical breakdown of MeHg in aquatic environments (Bergquist and Blum 2007; Tsui et al. 2020). The slope of Δ201Hg versus Δ199Hg is commonly used to evaluate whether inorganic mercury or MeHg photochemistry is driving the odd mass-independent fractionation (Tsui et al. 2020). For eiders in the study area, the slope of 1.42 ± 0.06 (Supplemental Fig. S5; linear regression R2 = 0.96, p < 0.001, n = 25) was close to the experimentally-derived slope of 1.36 ± 0.02 for fractionation due to MeHg photodemethylation (Bergquist and Blum 2007), indicating the higher Δ199Hg values of Chisasibi eiders (mean ± SD = 1.84 ± 0.42‰) were likely due to a source of more photo-degraded MeHg. The average Δ199Hg values of eiders from the other communities ranged from 0.86 to 1.26‰. However, three individual eiders from Umiujaq or Kuujjuarapik had higher Δ199Hg values, suggesting exposure to more highly photo-degraded MeHg was not limited to the Chisasibi area (Supplemental Fig. S5). Published data for estuarine and coastal marine fish in other geographic areas typically have lower Δ199Hg values between ∼0 and 1‰, which has been interpreted as minor photodemethylation of MeHg in coastal environments (Senn et al. 2010; Gehrke et al. 2011; Kwon et al. 2014; Rua-Ibarz et al. 2019; Tsui et al. 2020). The only published mercury isotope data for common eider (to our knowledge) are from estuarine food webs in the northeastern United states where Δ199Hg values of the birds (0.68–0.82‰) were similar though slightly lower than eiders from our sites in east Hudson Bay (Kwon et al. 2014). In contrast, the higher Δ199Hg values of Chisasibi eiders were comparable to freshwater fish of the Churchill River in subarctic eastern Canada (∼1–3‰), which had higher values than coastal marine fish downstream, presumably due to greater photodegradation of MeHg in the river system (Li et al. 2016).
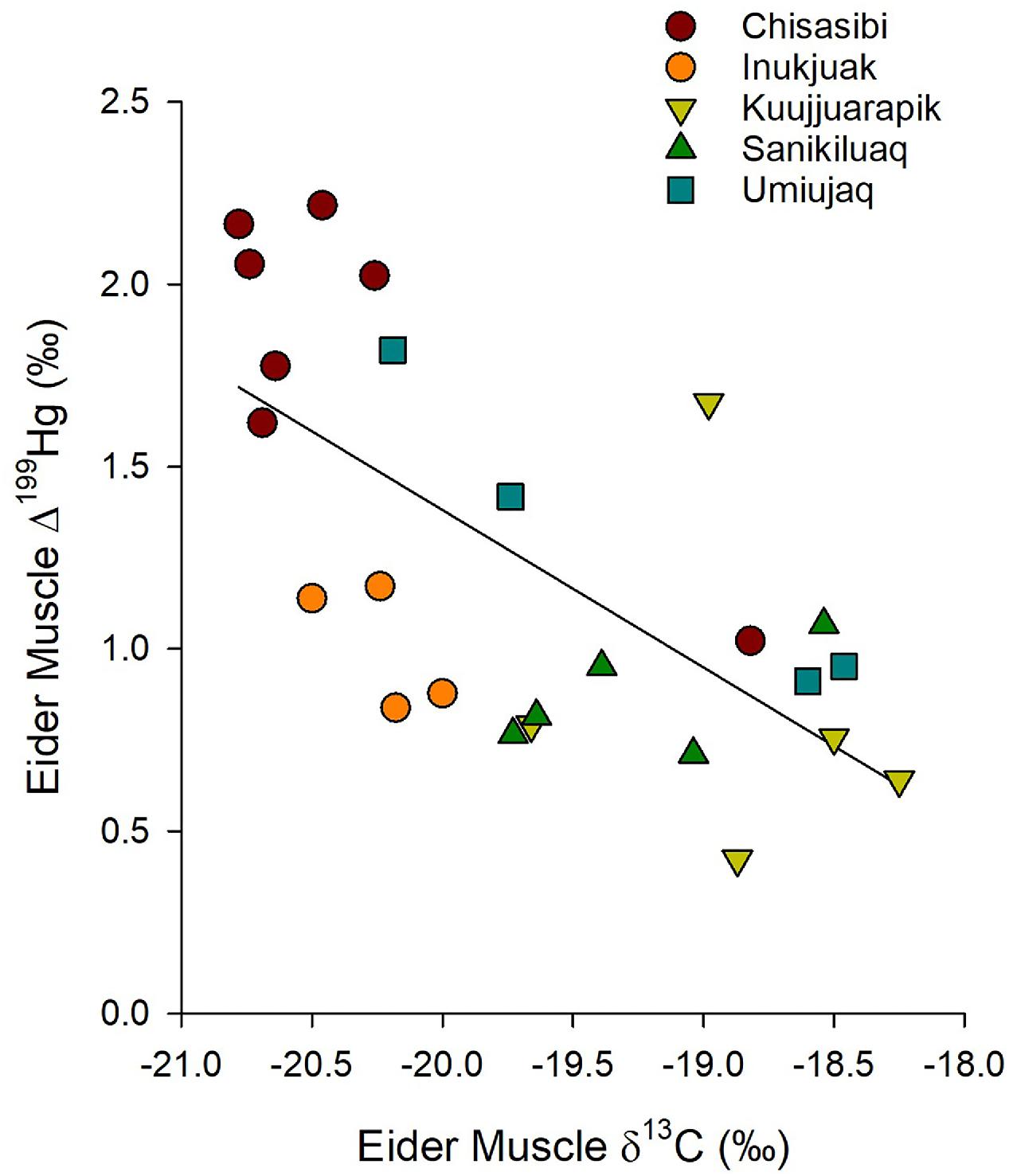
The mercury isotope ratios of eiders indicated greater mercury bioaccumulation was associated with a distinct exposure pathway. The THg concentration of eider muscle was positively associated with its Δ199Hg value; thus, eiders with higher mercury burdens were exposed to more highly photo-degraded MeHg (Fig. 7; linear regression R2 = 0.61, p < 0.001, n = 25). Further, a negative correlation between the δ13C and Δ199Hg values of eider muscle indicated that a diet supported by pelagic carbon led to greater exposure to photo-degraded MeHg (Fig. 8; linear regression R2 = 0.45, p < 0.001, n = 25). Together, these results suggest that a source of more highly photodegraded MeHg in the water column contributed to elevated mercury in the common eiders.
Fig. 7.
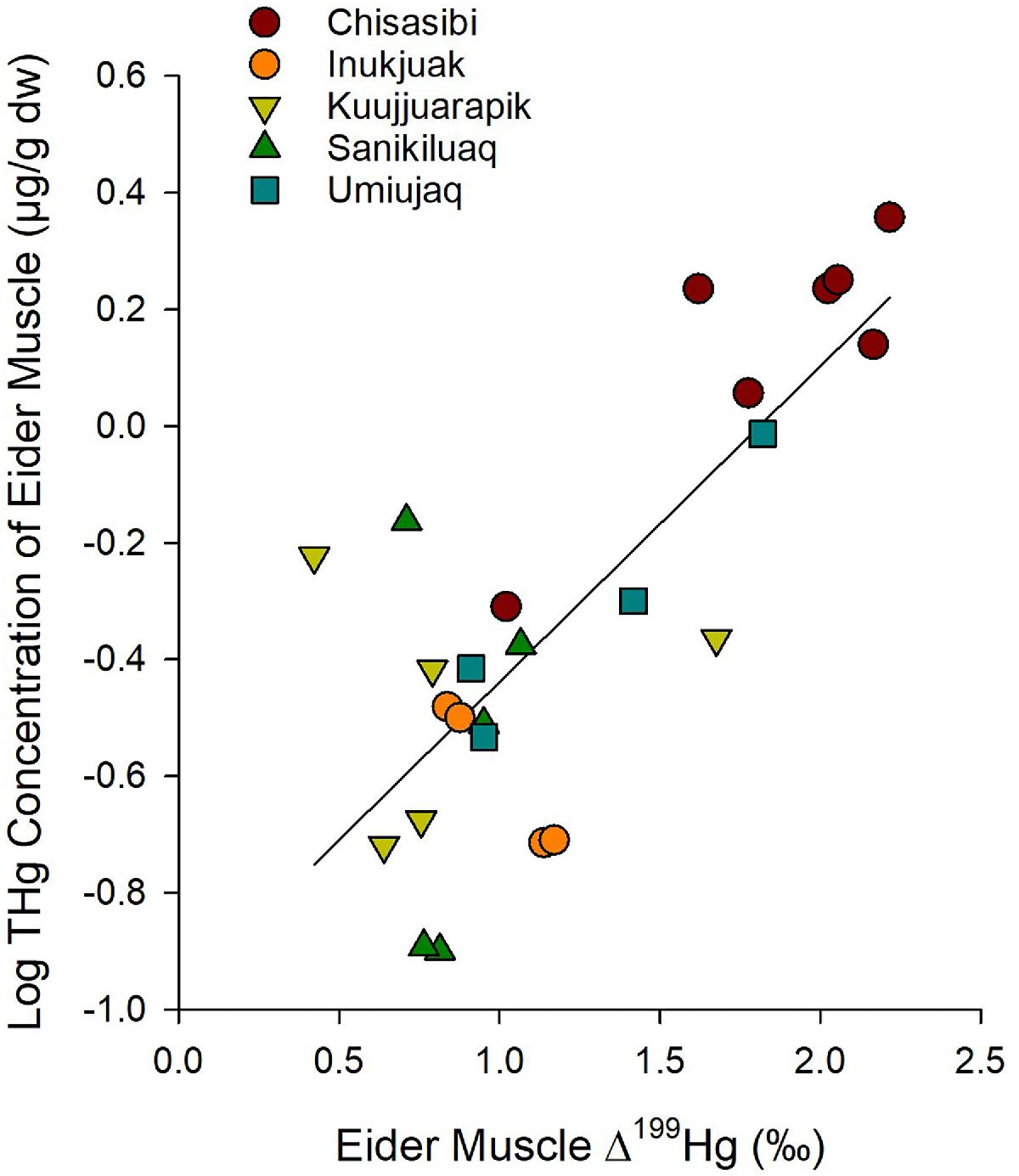
Fig. 8.
Source of methylmercury to the coastal food web in east James Bay
Multiple lines of evidence suggest elevated mercury in the coastal food web of east James Bay near Chisasibi was due to mercury loading from the La Grande River. First, blue mussels and common eiders were collected within the coastal plume of the La Grande River (Supplemental Fig. S6), a large system that discharges up to 5 000 m3/s of water to James Bay. The impact of this river discharge on seawater salinity is evident up to ∼100 km north along the coast during winter and ∼60 km during summer (Peck et al. 2022). Second, the mercury burden of blue mussels was positively associated with crustal elements (iron, vanadium, cobalt, and uranium), suggesting a link between river export and coastal accumulation. Finally, the mercury stable isotopes of eiders from James Bay near Chisasibi were distinct from eiders in Hudson Bay and the isotopic signature was consistent with photodegraded MeHg of freshwater origin (Li et al. 2016).
We hypothesize that riverine MeHg is entering the coastal food web via uptake by suspension feeding invertebrates consuming MeHg of freshwater origin. Methylmercury concentrations of water in the La Grande River (0.07–0.23 ng/L) are relatively low in comparison to other rivers draining into east James Bay (Fink‐Mercier et al. 2022). However, the La Grande River exports the largest annual load of MeHg to east James Bay, an estimated 5.79 kg/year (Fink‐Mercier et al. 2022), which likely settles along the coast due to flocculation in saline waters (Demarty et al. 2021). This annual MeHg export from the La Grande River is also much higher than estimates for major rivers in east Hudson Bay (Great Whale River and Innuksuac River) (Sonke et al. 2018). Methylmercury in water of the La Grande River is predominately in the dissolved fraction (Fink‐Mercier et al. 2022), and therefore transport of particulate MeHg is likely less important. In addition, associations of pelagic feeding (based on δ13C) with THg concentrations and Hg isotope values of eiders suggest that the mercury originated from the water column rather than sediment. Estuarine sediments can be sites of high MeHg production, though the relative importance of sediment versus water column sources of MeHg varies greatly among sites (Chen et al. 2014; Jonsson et al. 2014; Kwon et al. 2014; Taylor et al. 2019; Jonsson et al. 2022). It is possible that environmental characteristics of sediment in James Bay may be more amenable to MeHg production than in Hudson Bay. It is also possible that in situ water column methylation along the James Bay coast may be stimulated by inputs of terrestrial organic matter and nutrients, as observed in another boreal subarctic system (Schartup et al. 2015). Further research is needed to characterize coastal processes leading to enhanced mercury bioaccumulation in the marine food web of east James Bay. In addition, more detailed measurements are needed to determine the spatial extent of enhanced mercury bioaccumulation on the James Bay coast and the degree of exposure to other marine species connected to the pelagic food web.
HydroQuebec has implemented an extensive mercury monitoring program of the La Grande hydroelectric complex for decades (Bilodeau et al. 2017), though downstream effects were primarily considered in the context of a sequential system of reservoirs and the river. To our knowledge, few baseline data were collected for levels of MeHg in the coastal marine food web near Chisasibi during the initial construction and operation of reservoirs in the 1970s–1990s (Lucotte et al. 1999). It was assumed that the export of mercury to the James Bay coast was limited and localized (Schetagne and Verdon 1999), though pre-1996 data for fourhorn sculpin (Myoxocephalus quadricornis) collected within the coastal summer plume of the La Grande River had THg concentrations (0.31–0.55 µg/g wet weight, n = 106) that were several times higher than sculpin at sites outside the plume (0.10–0.22 µg/g wet weight, n = 60) (Schetagne and Verdon 1999). Beyond those observations, no long-term monitoring of the potential impact of reservoir discharge to the marine environment was developed.
Conclusion
This research found evidence for a river effect on mercury fate in a coastal subarctic food web based on investigation of two trophic levels and multiple stable isotope tracers. New community-driven study has since been initiated to further evaluate mercury levels in traditional foods from the coastal region of Eeyou Istchee near Chisasibi. Current conceptual modelling assumes that hydroelectric developments have little impact on the marine environment (Demarty et al. 2021). The findings from this study and other research in subarctic Canada (Kirk and Louis 2009; Schartup et al. 2015) suggest river export of mercury to northern coastal environments could impact the marine food web. The degree of influence is likely a function of the load of mercury entering the coastal environment. Beyond the study area, it remains unclear to what extent large rivers affect MeHg bioaccumulation on coastal shelves in the Arctic (Lehnherr 2014; Wang et al. 2018). The role of large rivers in exporting mercury to northern coasts and enhancing bioaccumulation in marine food webs is a critical link in the mercury cycle that requires further investigation.
Acknowledgements
This research was supported by funding from the Northern Contaminants Program (Crown-Indigenous Relations and Northern Affairs Canada, Project CB-06 and CB-08, 2013-2018) and Environment and Climate Change Canada (ECCC). Animal collections and transport were permitted under licences from the Canadian Wildlife Service (SC-55, SC-58), Fisheries and Oceans Canada (Quebec and Nunavut regions), and the Nunavut Department of Environment (2015-047, 2016-052, 2017-051). We gratefully acknowledge the community coordinators and hunters as a part of the Arctic Eider Society's Community-Driven Research Network from Chisasibi, Kuujjuarapik, Inukjuak, Sanikiluaq, and Umiujaq who collected marine animals for the study. This research would not have been possible without your contributions. We thank from Chisasibi: Vincent Gauthier-Doucet, Eddie Lameboy, Robert Lameboy, Jimmy Snowboy, and Christopher Sealhunter; from Inukjuak: Bobby Aculiak, Allie Aculiak, Billy Brian Kasudluak, Davidee Mina, Conilucy Naktialuk, and Peter Nulukie; from Kuujjuarapik: Charlie Angatookalook, Jimmy Paul Angatookalook, Jordan Kronenburg, George Tooktoo, Vincent Tooktoo, Samson Tooktoo, Eddy Tooktoo, and Willie Novalinga; from Sanikiluaq: Johnny Kudluarok, Stephan Kudluarok, Simeonie Uppik, Josie Amitook, and Salamonie Kowcharlie; and from Umiujaq: Johnny Kasudluak, Peter Crow, Charlie Kumarluk, Suzanna Aragutak, and Randy Aragutak. We also thank George Lameboy (Cree Nation of Chisasibi) and Annie Kasudluak Nalukturuk (Niqautik Hunters Association of Inukjuak) for their leadership during the study. We thank Michelle Zanuttig, Jian Jun Yang, Emily Porter, and Yueting Shao at the National Wildlife Research Centre (ECCC) for their technical assistance with chemical analyses. We gratefully acknowledge the production of the study area maps by Kuzey Gunesli and thank two reviewers for their constructive comments.
References
Ackerman J.T., Eagles-Smith C.A., Herzog M.P., Hartman C.A., Peterson S.H., Evers D.C., et al. 2016. Avian mercury exposure and toxicological risk across western North America: a synthesis. Science of the Total Environment, 568: 749–769.
Andersen M.B., Stirling C.H., Porcelli D., Halliday A.N., Andersson P.S., Baskaran M. 2007. The tracing of riverine U in Arctic seawater with very precise 234 U/238 U measurements. Earth and Planetary Science Letters, 259(1): 171–185.
Arctic Eider Society. 2024. Community-Driven Research Network. Website of the Arctic Eider Society, Sanikiluaq, NU. Available from https://arcticeider.com/community-driven-research-network/ [accessed February 2024].
Bergquist B.A., Blum J.D. 2007. Mass-dependent and -independent fractionation of Hg isotopes by photoreduction in aquatic systems. Science, 318(5849): 417–420.
Beyer J., Green N.W., Brooks S., Allan I.J., Ruus A., Gomes T., et al. 2017. Blue mussels (Mytilus edulis spp.) as sentinel organisms in coastal pollution monitoring: a review. Marine Environmental Research, 130: 338–365.
Bilodeau F., Therrien J., Schetagne R. 2017. Intensity and duration of effects of impoundment on mercury levels in fishes of hydroelectric reservoirs in northern Québec (Canada). Inland Waters, 7(4): 493–503.
Boyd P.W., Ellwood M.J. 2010. The biogeochemical cycle of iron in the ocean. Nature Geoscience, 3(10): 675–682.
Boyle E.A., Edmond J.M., Sholkovitz E.R. 1977. The mechanism of iron removal in estuaries. Geochimica et Cosmochimica Acta, 41(9): 1313–1324.
Braune B.M., Malone B.J., Burgess N.M., Elliott J.E., Garrity N., Hawkings J., et al. 1999. Chemical residues in waterfowl and gamebirds harvested in Canada. 1987–1995.
Braune B.M., Gaston A.J., Hobson K.A., Gilchrist H.G., Mallory M.L. 2014. Changes in food web structure alter trends of mercury uptake at two seabird colonies in the Canadian Arctic. Environmental Science & Technology, 48(22): 13246–13252.
Braune B., Chételat J., Amyot M., Brown T., Clayden M., Evans M., et al. 2015. Mercury in the marine environment of the Canadian Arctic: review of recent findings. Science of the Total Environment, 509-510: 67–90.
Brown T.M., Fisk A.T., Wang X., Ferguson S.H., Young B.G., Reimer K.J., et al. 2016. Mercury and cadmium in ringed seals in the Canadian Arctic: influence of location and diet. Science of the Total Environment, 545–546: 503–511.
Cardona‐Marek T., Knott K.K., Meyer B.E., O'Hara T.M. 2009. Mercury concentrations in southern Beaufort sea polar bears: variation based on stable isotopes of carbon and nitrogen. Environmental Toxicology and Chemistry, 28(7): 1416–1424.
Chastel O., Fort J., Ackerman J.T., Albert C., Angelier F., Basu N., et al. 2022. Mercury contamination and potential health risks to Arctic seabirds and shorebirds. Science of the Total Environment, 844.
Chen C.Y., Borsuk M.E., Bugge D.M., Hollweg T., Balcom P.H., Ward D.M., et al. 2014. Benthic and pelagic pathways of methylmercury bioaccumulation in estuarine food webs of the Northeast United States. PLoS ONE, 9(2): e89305.
Chételat J., Ackerman J.T., Eagles-Smith C.A., Hebert C.E. 2020. Methylmercury exposure in wildlife: a review of the ecological and physiological processes affecting contaminant concentrations and their interpretation. Science of the Total Environment, 711: 135117.
Cossa D., Tabard A.-M. 2020. Mercury in marine mussels from the St. Lawrence Estuary and Gulf (Canada): a mussel watch survey revisited after 40 years. Applied Sciences, 10(21): 7556.
Demarty M., Bilodeau F., Tremblay A. 2021. Mercury export from freshwater to estuary: carbocentric science elucidates the fate of a toxic compound in aquatic boreal environments. Frontiers in Environmental Science, 9.
Déry S.J., Mlynowski T.J., Hernández-Henríquez M.A., Straneo F. 2011. Interannual variability and interdecadal trends in Hudson Bay streamflow. Journal of Marine Systems, 88(3): 341–351.
Déry S.J., Stadnyk T.A., MacDonald M.K., Gauli-Sharma B. 2016. Recent trends and variability in river discharge across northern Canada. Hydrology and Earth System Sciences, 20(12): 4801–4818.
Dietz R., Fort J., Sonne C., Albert C., Bustnes J.O., Christensen T.K., et al. 2021. A risk assessment of the effects of mercury on Baltic Sea, Greater North Sea and North Atlantic wildlife, fish and bivalves. Environment International, 146: 106178.
Fink‐Mercier C., Lapierre J.-.F., Amyot M., del Giorgio P.A. 2022. Concentrations and yields of total Hg and MeHg in large boreal rivers linked to water and wetland coverage in the watersheds. JGR Biogeosciences, 127(5): e2022JG006892.
Gehrke G.E., Blum J.D., Slotton D.G., Greenfield B.K. 2011. Mercury isotopes link mercury in San Francisco Bay forage fish to surface sediments. Environmental Science & Technology, 45(4): 1264–1270.
Godin P., Macdonald R.W., Kuzyk Z.Z.A., Goñi M.A., Stern G.A. 2017. Organic matter compositions of rivers draining into Hudson Bay: present-day trends and potential as recorders of future climate change. JGR Biogeosciences, 122(7): 1848–1869.
Goudie R.I., Robertson G.J., A. R. 2020. Common eider (Somateria mollissima), version 1.0, In Birds of the world, Edited by S. M. Billerman. Cornell Lab of Ornithology, Ithaca, NY, USA.
Government of Canada. 2022. A East Hudson Bay Network research initiative on regional metal accumulation in the marine food web. Available from https://open.canada.ca/data/dataset/7efe1489-ca20-4553-804a-d66f67124a70 [accessed February 2024].
Hall B.D., St. Louis V.L., Rolfhus K.R., Bodaly R.A., Beaty K.G., Paterson M.J., et al. 2005. Impacts of reservoir creation on the biogeochemical cycling of methyl mercury and total mercury in boreal upland forests. Ecosystems, 8(3): 248–266.
Hamilton AL. 2011. The Hudson Bay complex in flux: contemplating the future of the world's largest seasonally ice-covered inland sea. International Institute for Sustainable Development, Winnipeg, Manitoba, Canada.
Hare A., Stern G.A., Macdonald R.W., Kuzyk Z.Z., Wang F. 2008. Contemporary and preindustrial mass budgets of mercury in the Hudson Bay marine system: the role of sediment recycling. Science of the Total Environment, 406(1–2): 190–204.
Hilgendag I.R., Swanson H.K., Lewis C.W., Ehrman A.D., Power M. 2022. Mercury biomagnification in benthic, pelagic, and benthopelagic food webs in an Arctic marine ecosystem. Science of the Total Environment, 156424.
Hudson Bay Consortium. 2020. The Hudson Bay Consortium 2020 Progress Report. Hudson Bay Consortium Secretariat.
HydroQuebec. 2023. Société d'énergie de la Baie James—the La Grande Complex. Available from http://www.hydroquebec.com/sebj/en/lagrande.html [accessed April 2023].
Jilbert T., Asmala E., Schröder C., Tiihonen R., Myllykangas J.P., Virtasalo J.J., et al. 2018. Impacts of flocculation on the distribution and diagenesis of iron in boreal estuarine sediments. Biogeosciences, 15(4): 1243–1271.
Jokinen S.A., Jilbert T., Tiihonen-Filppula R., Koho K. 2020. Terrestrial organic matter input drives sedimentary trace metal sequestration in a human-impacted boreal estuary. Science of the Total Environment, 717: 137047.
Jonsson S., Skyllberg U., Nilsson M.B., Lundberg E., Andersson A., Björn E. 2014. Differentiated availability of geochemical mercury pools controls methylmercury levels in estuarine sediment and biota. Nature Communications, 5.
Jonsson S., Liem-Nguyen V., Andersson A., Skyllberg U., Nilsson M.B., Lundberg E., et al. 2022. Geochemical and dietary drivers of mercury bioaccumulation in estuarine benthic invertebrates. Environmental Science & Technology, 56(14): 10141–10148.
Jörundsdóttir H.Ó., Jensen S., Hylland K., Holth T.F., Gunnlaugsdóttir H., Svavarsson J., et al. 2014. Pristine Arctic: background mapping of PAHs, PAH metabolites and inorganic trace elements in the North-Atlantic Arctic and sub-Arctic coastal environment. Science of the Total Environment, 493: 719–728.
Keller W., Paterson A., Rühland K., Blais J. 2014. Introduction-environmental change in the Hudson and James Bay region. Arctic, Antarctic, and Alpine Research, 46(1): 2–5.
Khoo C.L.L., Sipler R.E., Fudge A.R., Beheshti Foroutani M., Boyd S.G., Ziegler S.E. 2022. Salt-induced flocculation of dissolved organic matter and iron is controlled by their concentration and ratio in boreal coastal systems. JGR Biogeosciences, 127(11): e2022JG006844.
Kirk J.L., Louis V.L.S. 2009. Multiyear total and methyl mercury exports from two major sub-arctic rivers draining into Hudson Bay, Canada. Environmental Science & Technology, 43(7): 2254–2261.
Krachler R., Krachler R.F. 2021. Northern high-latitude organic soils as a vital source of river-borne dissolved iron to the ocean. Environmental Science & Technology, 55(14): 9672–9690.
Kwon S.Y., Blum J.D., Chen C.Y., Meattey D.E., Mason R.P. 2014. Mercury isotope study of sources and exposure pathways of methylmercury in estuarine food webs in the Northeastern U.S. Environmental Science & Technology, 48(17): 10089–10097.
Lehnherr I. 2014. Methylmercury biogeochemistry: a review with special reference to Arctic aquatic ecosystems. Environmental Reviews, 22(3): 229–243.
Lemire M., Kwan M., Laouan-Sidi A.E., Muckle G., Pirkle C., Ayotte P., et al. 2015. Local country food sources of methylmercury, selenium and omega-3 fatty acids in Nunavik, Northern Quebec. Science of the Total Environment, 509–510: 248–259.
Li M., Schartup A.T., Valberg A.P., Ewald J.D., Krabbenhoft D.P., Yin R., et al. 2016. Environmental origins of methylmercury accumulated in subarctic estuarine fish indicated by mercury stable isotopes. Environmental Science & Technology, 50(21): 11559–11568.
Lucotte M., Schetagne R., Thérien N., Langlois C., Tremblay A. 1999. Natural environments and hydroelectric reservoirs of northern Québec (Canada). Springer Berlin Heidelberg, Berlin, Heidelberg.
Mallory M.L., Braune B.M., Wayland M., Gilchrist H.G., Dickson D.L. 2004. Contaminants in common eiders (Somateria mollissima) of the Canadian Arctic. Environmental Reviews, 12(4): 197–218.
Mallory M.L., Loseto L.L., Ferguson S.H. 2010. The future of Hudson Bay: new directions and research needs. In A little less Arctic: top predators in the world's largest northern inland sea, Hudson Bay. pp. 291–303.
Mallory C.D., Gilchrist H.G., Robertson G.J., Provencher J.F., Braune B.M., Forbes M.R., et al. 2017. Hepatic trace element concentrations of breeding female common eiders across a latitudinal gradient in the eastern Canadian Arctic. Marine Pollution Bulletin, 124(1): 252–257.
McDonald M., Arraqutainaq L., Novalinga Z. 1997. Voices from the bay. Traditional ecological knowledge of Inuit and Cree in the Hudson Bay bioregion. Canadian Arctic Resources Committee,Ottawa.
Meattey D.E., Savoy L., Beuth J., Pau N., O'Brien K., Osenkowski J., et al. 2014. Elevated mercury levels in a wintering population of common eiders (Somateria mollissima) in the northeastern United States. Marine Pollution Bulletin, 86(1): 229–237.
Morris A.D., Braune B.M., Gamberg M., Stow J., O'Brien J., Letcher R.J. 2022. Temporal change and the influence of climate and weather factors on mercury concentrations in Hudson Bay polar bears, caribou, and seabird eggs. Environmental Research, 112169.
Murphy G.E.P., Dunic J.C., Adamczyk E.M., Bittick S.J., Côté I.M., Cristiani J., et al. 2021. From coast to coast to coast: ecology and management of seagrass ecosystems across Canada. Facets, 6(1): 139–179.
NTK. 2008. A life vest for Hudson Bay's drifting stewardship. Arctic, 61(SUPPL. 1): 35–47.
Ouellet J.-.F., Vanpé C., Guillemette M. 2013. The body size-dependent diet composition of North American sea ducks in winter. PLoS ONE, 8(6): e65667.
Peck C.J., Kuzyk Z.Z.A., Heath J.P., Lameboy J., Ehn J.K. 2022. Under-ice hydrography of the La Grande River plume in relation to a ten-fold increase in wintertime discharge. JGR Oceans, 127(10): e2021JC018341.
Porvari P., Verta M. 1995. Methylmercury production in flooded soils: a laboratory study. Water, Air, & Soil Pollution, 80(1–4): 765–773.
Provencher J.F., Gilchrist H.G., Mallory M.L., Mitchell G.W., Forbes M.R. 2016. Direct and indirect causes of sex differences in mercury concentrations and parasitic infections in a marine bird. Science of the Total Environment, 551–552: 506–512.
R Core Team. 2021. R: a language and environment for statistical computing. R Foundation for Statistical Computing. Available from https://www.R-project.org/.
Rohonczy J., Chételat J., Robinson S.A., Arraqutainaq L., Heath J.P., McClelland C., et al. 2024. Contrasting trophic transfer patterns of cadmium and mercury in the Arctic marine food web of east Hudson Bay, Canada. Environmental Science and Pollution Research. In press.
Rosa M., Ward J.E., Shumway S.E. 2018. Selective capture and ingestion of particles by suspension-feeding bivalve molluscs: a review. Journal of Shellfish Research, 37(4): 727–746.
Rua-Ibarz A., Bolea-Fernandez E., Maage A., Frantzen S., Sanden M., Vanhaecke F. 2019. Tracing mercury pollution along the Norwegian coast via elemental, speciation, and isotopic analysis of liver and muscle tissue of deep-water marine fish (Brosme brosme). Environmental Science & Technology, 53(4): 1776–1785.
Saniewska D., Bełdowska M., Szymczak E., Kuliński K., Bełdowski J., Voss M., et al. 2022. Processes affecting the transformation of mercury in the coastal zone in the vicinity of two river mouths in the southern Baltic Sea. Marine Chemistry, 238: 104065.
Schartup A.T., Balcom P.H., Soerensen A.L., Gosnell K.J., Calder R.S.D., Mason R.P., et al. 2015. Freshwater discharges drive high levels of methylmercury in Arctic marine biota. Proceedings of the National Academy of Sciences USA, 112(38): 11789–11794.
Schetagne R., Verdon R. 1999. Post-impoundment evolution of fish mercury levels at the la grande complex, Québec, Canada (from 1978 to 1996). In Mercury in the biogeochemical cycle: natural environments and hydroelectric reservoirs of Northern Québec (Canada). Edited by M. Lucotte, R. Schetagne, N. Thérien, C. Langlois, A. Tremblay. Springer Berlin Heidelberg, Berlin, Heidelberg. pp. 235–258.
Schlesinger W.H., Klein E.M., Vengosh A. 2017. Global biogeochemical cycle of vanadium. Proceedings of the National Academy of Sciences USA, 114(52): E11092–E11100.
Senn D.B., Chesney E.J., Blum J.D., Bank M.S., Maage A., Shine J.P. 2010. Stable isotope (N, C, Hg) study of methylmercury sources and trophic transfer in the Northern Gulf of Mexico. Environmental Science & Technology, 44(5): 1630–1637.
Soerensen A.L., Schartup A.T., Gustafsson E., Gustafsson B.G., Undeman E., Björn E. 2016. Eutrophication increases phytoplankton methylmercury concentrations in a coastal sea—a Baltic Sea case study. Environmental Science & Technology, 50(21): 11787–11796.
Sonke J.E., Teisserenc R., Heimbürger-Boavida L.E., Petrova M.V., Marusczak N., Le Dantec T., et al. 2018. Eurasian river spring flood observations support net Arctic Ocean mercury export to the atmosphere and Atlantic Ocean. Proceedings of the National Academy of Sciences USA, 115(50): E11586–E11594.
St. Louis V.L., Derocher A.E., Stirling I., Graydon J.A., Lee C., Jocksch E., et al. 2011. Differences in mercury bioaccumulation between polar bears (ursus maritimus) from the Canadian high- and sub-arctic. Environmental Science & Technology, 45(14): 5922–5928.
Stewart D.B., Barber D.G. 2010. The ocean-sea ice-atmosphere system of the Hudson Bay complex. In A little less Arctic: top predators in the world's largest northern inland sea, Hudson Bay. pp. 1–37.
Stewart D.B., Lockhart W.L. 2005. Summary of the Hudson Bay marine ecosystem overview. Canada Department of Fisheries and Oceans, Winnipeg, MB. vi + 66p.
Swanner E.D., Planavsky N.J., Lalonde S.V., Robbins L.J., Bekker A., Rouxel O.J., et al. 2014. Cobalt and marine redox evolution. Earth and Planetary Science Letters, 390: 253–263.
Taylor V.F., Buckman K.L., Seelen E.A., Mazrui N.M., Balcom P.H., Mason R.P., et al. 2019. Organic carbon content drives methylmercury levels in the water column and in estuarine food webs across latitudes in the Northeast United States. Environmental Pollution, 246: 639–649.
Tsui M.T.-.K., Blum J.D., Kwon S.Y. 2020. Review of stable mercury isotopes in ecology and biogeochemistry. Science of the Total Environment, 716: 135386.
Wang K., Munson K.M., Beaupré-Laperrière A., Mucci A., Macdonald R.W., Wang F. 2018. Subsurface seawater methylmercury maximum explains biotic mercury concentrations in the Canadian Arctic. Scientific Reports, 8(1).
Zhang Y., Jacob D.J., Dutkiewicz S., Amos H.M., Long M.S., Sunderland E.M. 2015. Biogeochemical drivers of the fate of riverine mercury discharged to the global and Arctic oceans. Global Biogeochemical Cycles, 29(6): 854–864.
Supplementary material
Supplementary Material 1 (DOCX / 1.19 MB).
- Download
- 1.19 MB
Information & Authors
Information
Published In

FACETS
Volume 9 • January 2024
Pages: 1 - 13
Editor: John P. Smol
History
Received: 13 July 2023
Accepted: 27 November 2023
Version of record online: 4 April 2024
Copyright
© 2024 Authors Heath, Arragutainaq, Lameboy, Mickpegak, and The Crown. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Data generated or analyzed during this study are available in the Government of Canada Open Data Portal, https://open.canada.ca/data/dataset/7efe1489-ca20-4553-804a-d66f67124a70.
Key Words
Sections
Subjects
Authors
Author Contributions
Conceptualization: JC, JPH, LA, RM
Data curation: CM
Formal analysis: JC
Funding acquisition: JC, JPH, LA, RM
Investigation: LA, JL, CM, RM
Project administration: JC, JPH, LA, JL, RM
Writing – original draft: JC
Writing – review & editing: JC, JPH, CM
Competing Interests
The authors declare there are no competing interests.
Funding Information
Northern Contaminants Program: 2013-2018, CB-06, CB-08
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
John Chételat, Joel P. Heath, Lucassie Arragutainaq, John Lameboy, Christine McClelland, and Raymond Mickpegak. 2024. River influence on mercury bioaccumulation in the coastal food web of Eeyou Istchee, James Bay, Canada. FACETS.
9(): 1-13. https://doi.org/10.1139/facets-2023-0121
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item