Our results show that gonadal investment in cod off Newfoundland is highly variable, but shows significant long-term patterns. For male cod, our findings agree with the prediction of life history theory that reproductive investment will increase during periods of high fishing mortality, and that reduced fishing mortality will cause such a trend to level off, eventually reversing it. Surprisingly, we do not find a similar pattern for female cod, despite our prior expectation that females, because of their more direct link between gonadal investment and fitness, should show a stronger pattern than males. This expectation should have applied especially to a species with a lek-like mating system, which is prone to reduce sperm competition among courting males compared with communal spawning. Before discussing possible causes of this difference, we first discuss other drivers of variability in gonad weight.
Interpretation of the effects of explanatory variables other than sampling year
The single most important variable explaining variability in gonad weight is, as expected, body weight: larger fish have larger gonads. According to our findings, this relationship is isometric only for females in one of the three studied populations. For females in the other two populations and for males in all three populations, the relationship shows positive allometry, so that GSI increases with body weight. We are not aware of earlier studies documenting such positive allometry in male cod, although it has been demonstrated in other species (
Stoltz et al. 2005).
Because the primary drivers of variation in gonad weight, beyond the effect of body weight, are poorly known in the cod populations studied here, we have tested an array of candidate explanatory variables, using standard model selection techniques to find the “best” model for each combination of sex, stock, and time span. When reporting on the resulting 12 models, there is an increased risk of a significant effect appearing by chance alone. We have reduced this risk of spurious effects by testing only variables and interactions that we considered biologically reasonable a priori, and when in doubt, we have checked that our main conclusions do not depend on including potentially spurious effects.
Among individual-level explanatory variables, the best models often include the relative liver index, which shows a positive relationship with gonad weight. This result is biologically expected and in agreement with earlier studies by
Yoneda and Wright (2004), who reported a positive correlation between the relative liver index and the potential fecundity of female cod for various stocks off Scotland, and by
Skjæraasen et al. (2006), who found a weak positive relationship between potential fecundity and lipid content at the time of spawning in cod (in cod, lipids are mainly stored in the liver;
Kjesbu et al. 1991;
Lambert and Dutil 1997). The influence of the relative liver index turns out to be relatively weak in our study as well. Similarly,
McIntyre and Hutchings (2003) concluded that, after accounting for the effect of body weight, accounting for the effect of the body condition index did not significantly improve the description of the potential fecundity of cod.
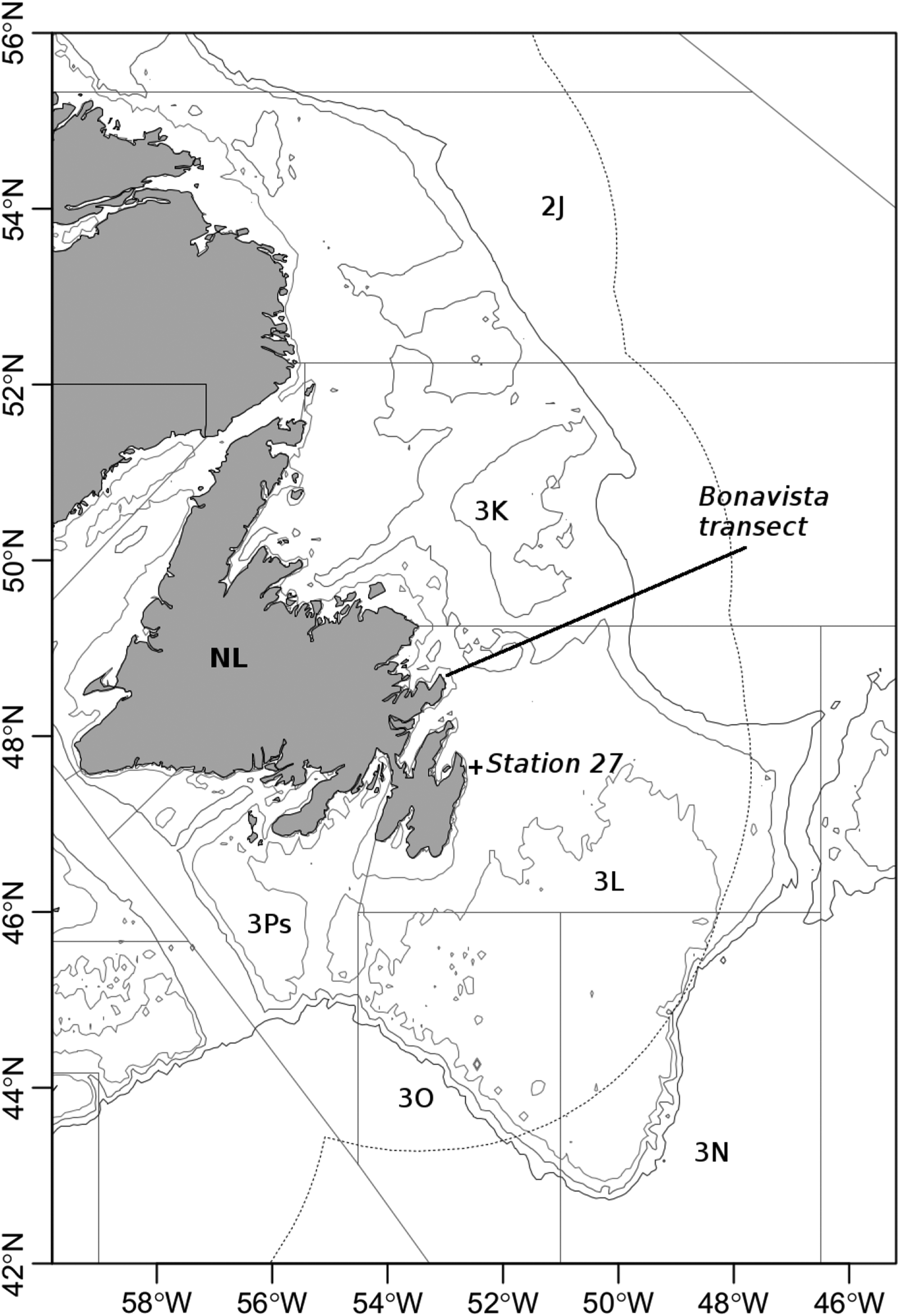
Among other significant explanatory variables, those accounting for sampling day (day of the year) and sampling location (longitude and latitude) frequently explain some variability in gonadal investment (with sampling day and longitude accounting for more variability than latitude). As sampling date and sampling location are only weakly correlated, we consider it likely that these statistically estimated effects are real biological effects. The increase in gonad weight throughout the season, accounted for in all our 12 models via the inclusion of the sampling day, illustrates the variability in gonad weight within the considered maturity stages. A significant geographical gradient could reflect a movement of individuals in the most advanced maturity stages toward spawning sites. This is in line with earlier reports of the geographic distribution of cod individuals depending on their maturity stage (
Morgan and Trippel 1996;
Lawson and Rose 2000;
Robichaud and Rose 2003). These studies, together with direct tank observations (
Hutchings et al. 1999), suggest the existence of a lek-like mating system in the studied cod populations, with male cod in spawning condition found on the continental shelf together with spawning females, whereas females with premature ovaries and spent females are more abundant on the continental slope.
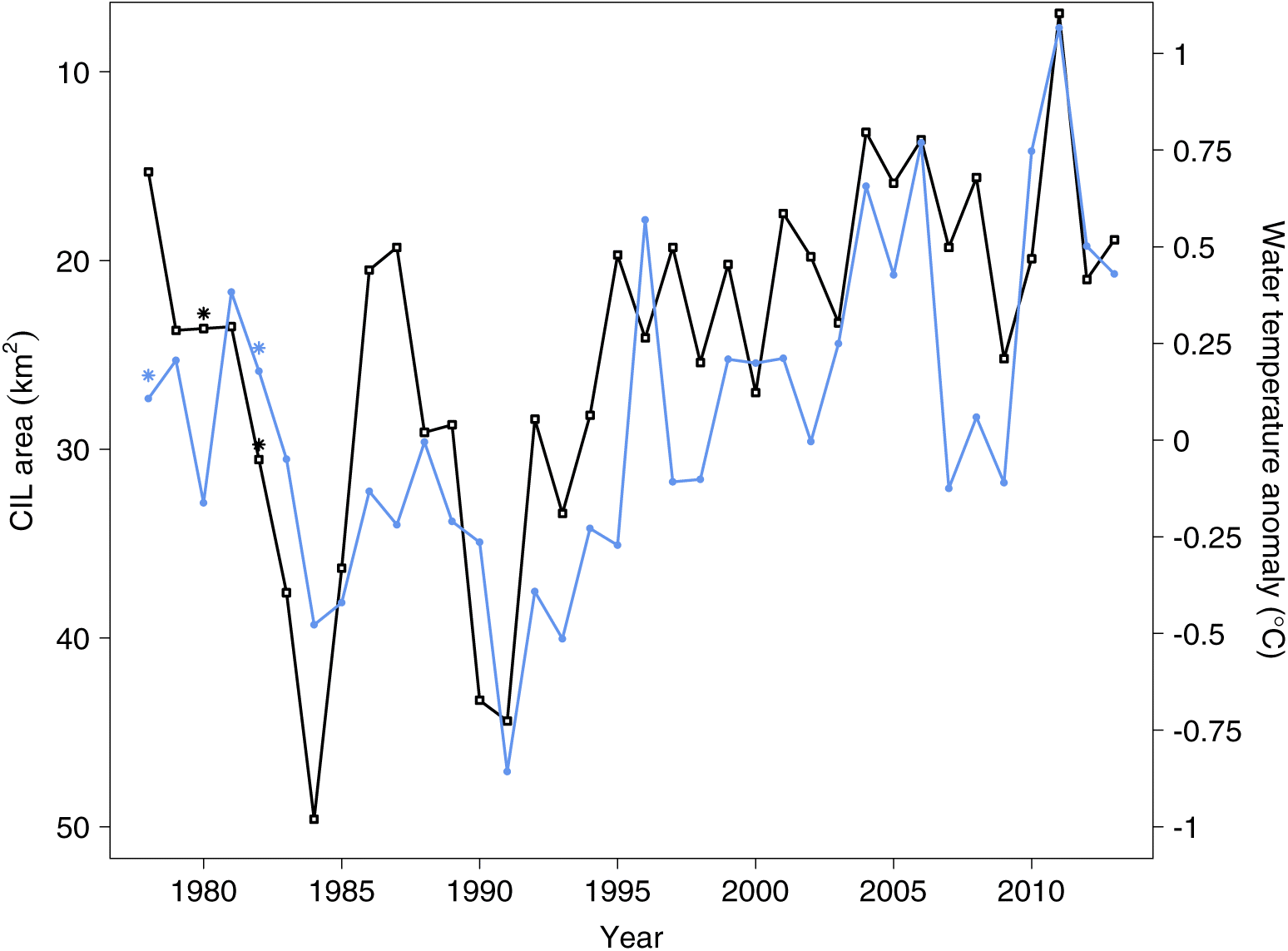
Interpreting the relationships between gonad weight and water temperature anomaly (showing a positive correlation when present) and between gonad weight and the area of the vertical cross section of the CIL (showing positive or negative correlations when present) requires some care. The CIL area and water temperature anomaly are negatively correlated (Pearson’s correlation coefficient
r = −0.70 for the years 1978–2013), and they never occur together in any of the 12 best models. Although the measurements underlying water temperature anomaly and CIL area are partly taken in Division 3L and are, thus, thought to reflect conditions in this division, they are also thought to be representative of the prevailing environmental conditions in Divisions 3NO, but less so those in Subdivision 3Ps. The estimated effect on gonad weight could reflect direct effects of temperature on physiological rates, or effects of oceanographic conditions on cod distribution and food availability. A positive effect of water temperature anomaly and a negative effect of CIL area on reproductive investment seem reasonable for populations living near the cold end of the species’ range (
Kjesbu et al. 1998;
Yoneda and Wright 2005;
Wright et al. 2011). Moreover,
Pinhorn (1984) proposed the influence of the water temperature as a potential cause of differences in fecundity observed among the stocks considered here. However, in Subdivision 3Ps, CIL area has a positive effect on the gonadal investment of females, which suggests a different mechanism linking oceanographic conditions and reproductive investment. For example,
Hutchings and Myers (1994) found a positive effect of cold water on spawning time and attributed this to cold on-shelf conditions in the area forcing the fish to stay in deeper warmer waters. Water temperature anomaly and CIL area do not have a large influence on our results. Importantly, disregarding these variables does not affect the significance of the trends reported in this study except for females in Subdivision 3Ps, where a significant negative trend in gonad weight appears for the full time span. In this sense, the consideration and potential inclusion of temperature effects in our models is conservative.
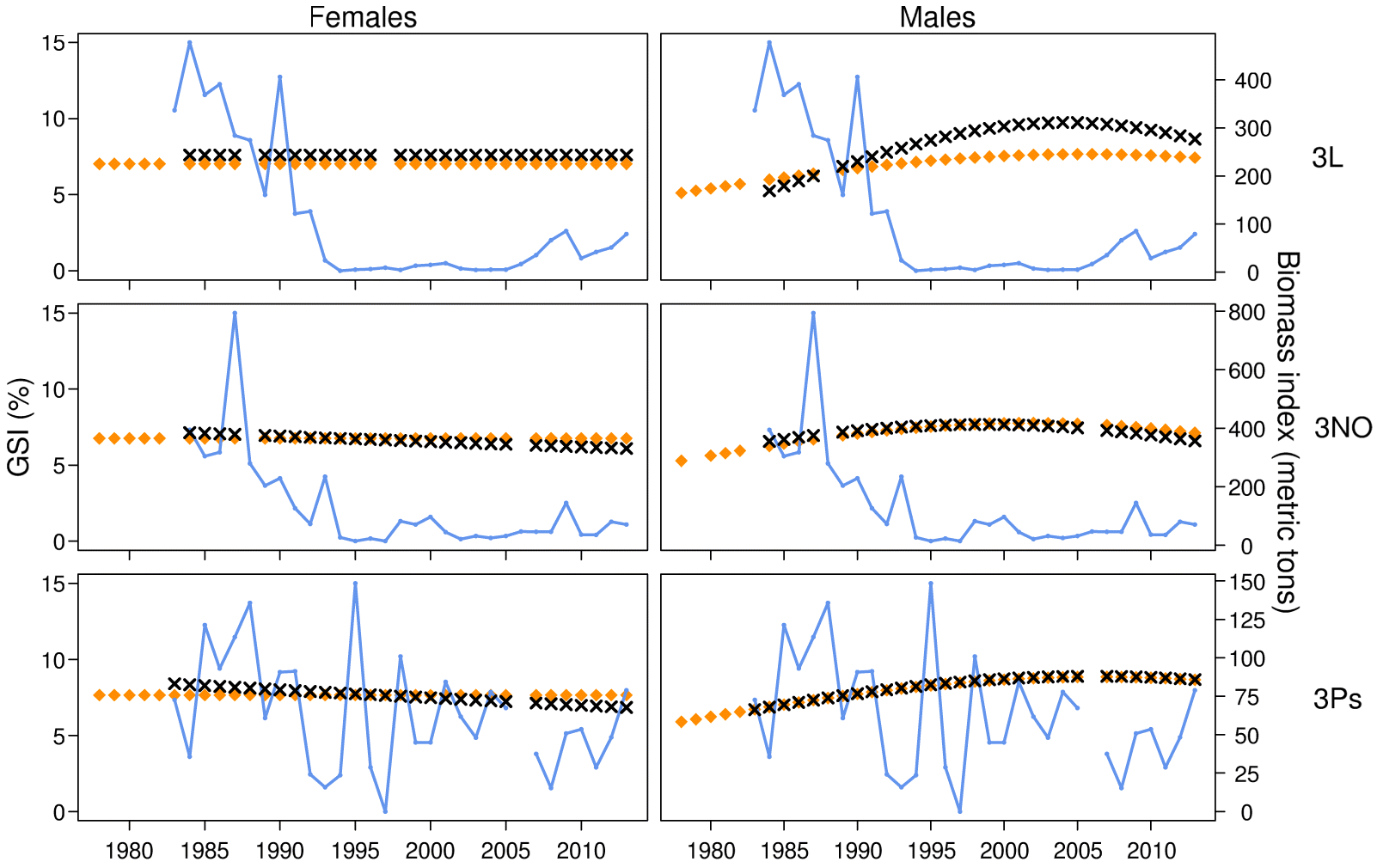
Biomass does not seem to be a key factor influencing gonad weight in the studied cod populations. The biomass index is retained in models for both sexes in Divisions 3NO and for females only in Subdivision 3Ps; when present, its influence on gonad weight is consistently negative. A negative effect of population density on fecundity has been shown in many fishes (e.g.,
Kipling and Frost 1969;
Healey 1978;
Koslow et al. 1995), including the cod stocks studied here (
Stares et al. 2007). As a robustness check, we have confirmed that the choice between considering the stock biomass either in the sampling year or in the previous year (when cod start to divert energy to the maturation of gametes to be released during the next spawning event) does not modify our conclusions about time trends, except for males in Divisions 3NO, where the latter choice suppresses the linear trend.
Of course, a stock biomass index is only a proxy for the per capita availability of resources and does not capture changes originating from the overall availability of resources. For example, the population size and availability of capelin are known to be important for the growth and reproduction of northern cod (
Rose and O’Driscoll 2002), but quantifying this interaction over long time periods would be challenging. Nevertheless, because the influence of stock biomass on fecundity—or, in our study, gonad weight—is mediated through the nutritional status of individuals, we may expect this influence to be stronger when the energetic condition of individuals (e.g., through the relative liver index) is not accounted for. In agreement with this expectation, the effect of biomass on gonad weight becomes significant in one case (females in Division 3L) when the relative liver index is excluded from the model. In some other cases, however, the biomass effects on gonad weight remain significant even when the relative liver index is included, which is indicative of a more complex link between stock biomass and gonad weight.
Even though some interannual variability has been reported in the spawning time of cod (
Hutchings and Myers 1994;
Morgan et al. 2013), this variability seems to be captured by the various individual characteristics (e.g., relative liver index) or sampling characteristics (e.g., longitude) we have considered. As a consequence, the interaction between sampling day and CIL area or water temperature anomaly appears not to bring any additional information, and hence, has never been retained in any of the 12 best models.
Interpretation of temporal trends in gonad weight
Our results reveal positive residual long-term trends in the gonadal investment of male cod in each of the three populations after the effects of individual, sampling-related, and environmental explanatory variables have been accounted for. Although we cannot rule out the possibility of unaccounted variables acting on phenotypic plasticity, the positive trends observed in males at least during the first halves of the time series are in accordance with the hypothesis that the period of heavy fishing has driven the evolution of higher reproductive investment.
A traditional assessment of the reproductive biology of cod suggests that males should be less likely to show evolutionary responses in gonadal investment than females. In a fish species like cod, featuring female mate choice (
Hutchings et al. 1999;
Nordeide and Folstad 2000;
Rowe et al. 2007), male reproductive success should depend less strongly on the number of produced gametes than female reproductive success. In other words, the importance of gonad weight in the determination of fitness is expected to be lower for males than for females. In support of this rationale,
Rakitin et al. (2001) did not find a significant relationship between reproductive success and the weight loss of male cod during spawning under experimental conditions. However, GSI values are similar between the two sexes (
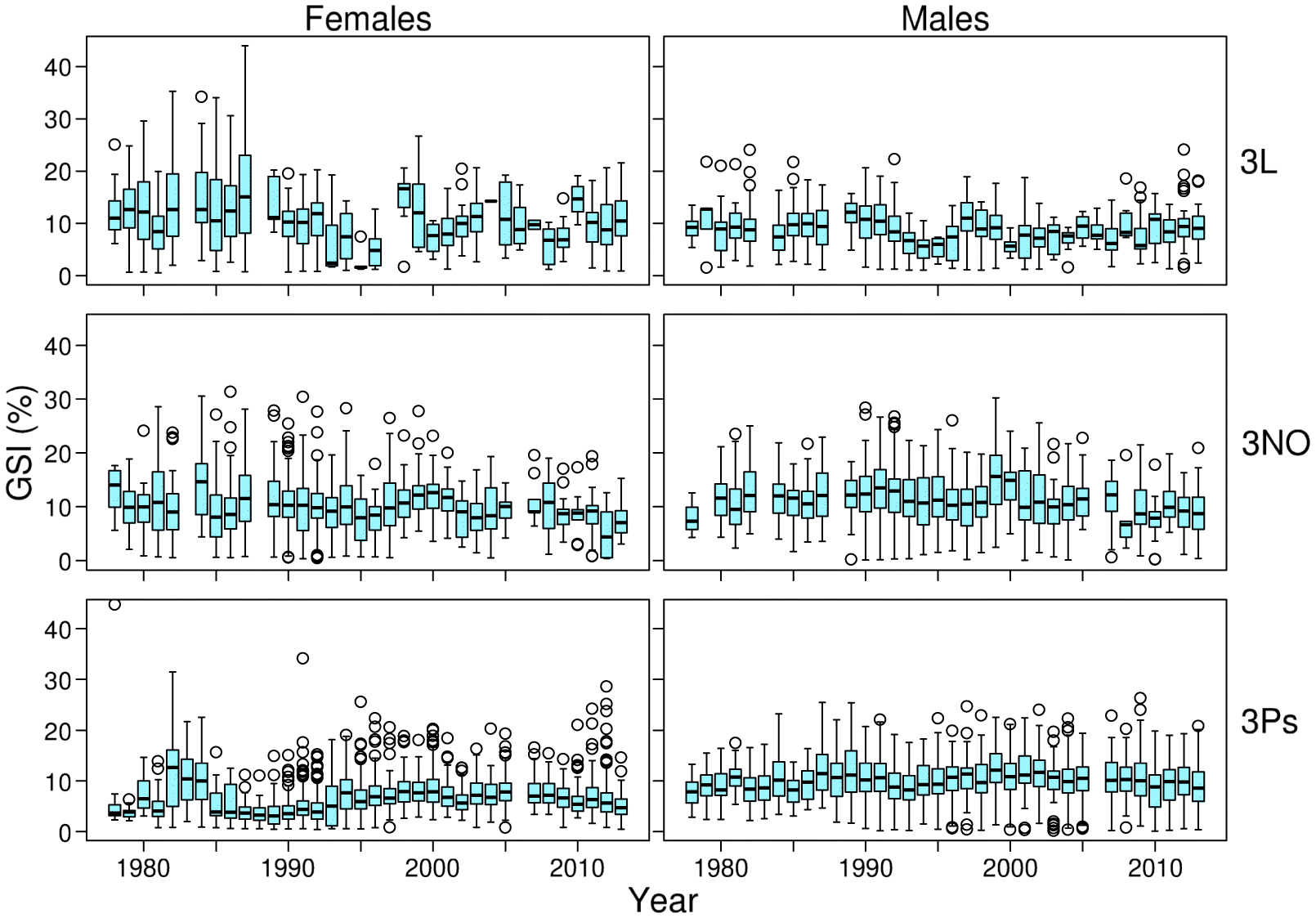
Fig. 4;
Burton et al. 1997), suggesting that an investment in large gonads is also important for male cod. Indeed, the production of higher numbers of spermatozoa can confer an advantage, as male cod mate with several females over the course of a spawning season (
Hutchings et al. 1999;
Bekkevold et al. 2002). Given that cod sperm remains viable for long periods in water, this may allow male cod to fertilize unfertilized eggs from other mating pairs (
Trippel and Morgan 1994).
Our analyses have provided no support for the hypothesis that increased mortality rates have caused the evolution of increased gonadal investment in female cod. However, there are several reasons why an actual evolutionary change might not be detected by our analyses, i.e., why they might be prone to false negatives (type-II errors). First, our methodology is conservative, in that we tested for the significance of the effect of sampling year only after all other significant covariates had been accounted for; specifically, we used a likelihood-ratio test to evaluate whether the inclusion of sampling year could improve a model in which several other significant explanatory variables were already present.
A second explanation for the absence of evidence for an evolutionary increase in female gonadal investment stems from the massive plasticity associated with this metric in cod, as already shown by
McIntyre and Hutchings (2003). Naturally, the high variability in gonad weight leads to a reduced statistical power of the analyses. We have tried to minimize this problem by focusing on a single maturity stage for females. Oocytes go through significant changes that influence egg diameter during the late maturity stages (
Kjesbu et al. 1990;
Kjesbu 1991), and the incorporation of proteins into the oocytes during these stages leads to an increase in gonad weight (
Eliassen and Vahl 1982). GSI is not static in males either, but even though male GSI displays a strong increase in the course of spermatogenesis (
Dahle et al. 2003), the production of spermatozoa takes place several months prior to spawning (
Burton et al. 1997;
Rideout and Burton 2000), leading to the relative stability of male GSI values during the maturity stage considered here (
Almeida et al. 2008). Indeed, our models for females explain a systematically lower portion of deviance and have higher standard errors for estimates of the effect of sampling year than our models for males. Nevertheless, the point estimates for linear time trends are consistently negative for females (
Table 1), suggesting that the absence of positive trends is genuine, rather than representing false negatives (type-II errors).
The prediction that fishing favors increased reproductive investment is qualitatively well established. Only recently, however, have quantitative predictions from eco-genetic models, parameterized for female life histories in cod-like species, shown that fisheries-induced selection not only favors increased GSI, but also that the resultant GSI evolution is slow relative to maturation evolution (
Dunlop et al. 2009;
Enberg et al. 2009; but see
Andersen and Brander 2009, who reached divergent conclusions). It is, therefore, possible that the time period during which two indispensable conditions hold jointly in the three studied populations—that we have adequate quantitative observations and that fisheries-induced selection was strong—has been too short (up to 20 years, and less for analyses including stock biomass) to cause detectable evolutionary change. Moreover, since the exploitation of the studied cod populations had already intensified in the 1950s (
Lilly et al. 2008), it is possible that adaptations to high mortality rates occurred before the time period covered in this study. Yet, we argue that adaptation to fishing is unlikely to have halted just a few decades after the industrialization of the Newfoundland cod fishery.
Our results show that changes in gonadal investment in males have leveled off or reversed in all studied stocks toward the end of the study period. This is in agreement with the prediction that a reduction in total mortality should reverse the evolution of reproductive investment, albeit at a slower pace (
Dunlop et al. 2009;
Enberg et al. 2009). Such a reversal in gonadal investment should become apparent only with a delay of several years, because it becomes expressed first when the progeny of selected parents reach maturity. The reported negative trends in the gonadal investment of females could also represent such an evolutionary reversal. However, these potential reversals should be interpreted with caution, as their degree is difficult to predict for the stocks studied here, for at least two reasons. First, even after the directed cod fisheries were closed, natural mortality may have remained high (
Shelton et al. 2006;
Lilly et al. 2013), and selection for increased reproductive investment may, thus, have persisted, albeit at reduced intensity. Second, some cod fishing has taken place during the moratoria in all stocks, and the moratorium for St. Pierre Bank stock was lifted in 1997 (
DFO 2015).
In summary, the results of the analyses performed here show evidence for an evolutionary change in gonadal investment for cod populations off Newfoundland, but only for male cod. This suggests that gonad weight has greater importance for the reproductive success of male cod than has previously been acknowledged. The lack of a similar response in female cod could be explained by the relatively short time span with strong fisheries selection, in conjunction with the higher phenotypic variability in gonadal investment in females compared with males; but this does not explain why the available evidence points to negative trends in females, if any. Our findings highlight that investigating the ecological and evolutionary determinants of reproductive investment in wild fish is not straightforward, and underscore how little we know about the reproduction of wild cod in particular, thus calling for more studies unraveling the reproductive biology of this important species.