Can fisheries be “regenerative”? Adapting agroecological concepts for fisheries and the blue economy
Abstract
Regenerative design, in which agricultural practices are organized to work with nutrient cycles and successional processes, is increasingly being explored in food systems research and practice. In this commentary, I explore whether regenerative design concepts can be adapted to marine contexts, given increased global interest in the potential of marine ecosystems to support sustainable development, i.e., the blue economy. There are numerous fundamental ecological differences between terrestrial and marine ecologies that make it difficult to directly translate regenerative farming's focus on managing the nutrient cycle. However, building on a framework for regenerative food systems that focuses on how production activities are organized rather than the specific practices and technologies in use, I find multiple useful parallels to familiar patterns in the fisheries literature, specifically, fishing down the food web, poverty traps, and portfolio-based fishing. I conclude with a discussion of directions for research on regenerative fisheries and concerns regarding the potential for greenwashing under the banner of a regenerative blue economy.
Introduction
Discussions about the future of terrestrial and marine food systems are rapidly shifting in tenor, moving from the now well-worn language of sustainability to arguably less anthropocentric framings that centre nature as much as people, e.g., regenerative and nature-positive solutions (Gaupp et al. 2021; Loring 2021). In terrestrial food systems research, regenerative agriculture—a form of agroecology that seeks to improve soil health and increase biologic carbon sequestration while also producing diverse and nutritious foods—is gaining ground as an alternative framing to sustainable agriculture (White 2020; Loring 2021). In the world of fisheries, there is similarly burgeoning interest in the notion of a “blue economy” that can be built around practices that provide nutritious and climate friendly food while restoring marine ecosystems and capturing and sequestering carbon in the ocean (Lovelock and Duarte 2019; Gentry et al. 2020; Koehn et al. 2022).
Many wild-caught fisheries are maximally fished or overfished, however, so the increasing global demand for aquatic foods that the blue economy narrative represents evokes multiple concerns given the depleted and arguably tenuous state of marine biodiversity (Costello et al. 2016; FAO 2022). For the last three decades, growth in the aquaculture sector has comprised almost all of the growth seen in global consumption of aquatic foods (FAO 2022), and numerous matters related to the impacts of aquaculture on marine ecosystems and the societal promises it can fulfil remain hotly debated (Barrett et al. 2019; Farmery et al. 2021). And, while it is true that many wild-caught fisheries have established reputations for sustainability, recent developments and surprise population declines in high-profile fisheries are bringing this dogma into question (Palomares et al. 2020; Cheney 2021). Even American lobster fisheries, which have long been considered a sustainability success story (Acheson 1990), are now being reconsidered because traditional practices in the fishery have negatively impacted ecosystem structure and species of concern (Steneck et al. 2011; Jacquet and Pauly 2022).
In this paper, I explore whether a similar turn toward the language of regeneration is possible and advisable for marine fisheries and other ocean-based foods. There are numerous fundamental ecological differences between terrestrial and marine ecologies, so it is important to explore whether regeneration as presently understood in terrestrial food systems sensibly translates to a marine setting. Below, I introduce the concepts and science behind the regenerative movement in agriculture and then argue that, despite many differences and with some appropriate caveats and adjustments, a regenerative systems framing can contribute to how we think about the future of wild fisheries and aquaculture. Further, I argue that this framing can also help address critical oversights in current ways of thinking about sustainability and the blue economy in general.
Regeneration
In general, regenerative design seeks to work with or otherwise mimic slow and fast cycles of ecological change, such as nutrient cycling and landscape succession (Pearson 2007; Rhodes 2017). Much regenerative agriculture focuses largely on soil and its role in the terrestrial nutrient cycle, that is, the movement and exchange of inorganic and organic matter into the production of new matter. In practice, regenerative agriculture often entails foregoing the use of chemical fertilizers and tillage in favour of such practices as crop rotation, fallowing, the use of nitrogen-fixing plants, perennial plants, livestock, and minimized soil disturbance (White 2020). Farmers implement these various practices in a multi-year strategy with the goal of building soil nutrients, structure, and microbial communities and sequestering carbon in the soil and in plant matter (Newton et al. 2020). In addition to the focus on soil, regenerative practices also emphasize landscape-level biodiversity, foregoing the use of chemicals for pest and weed control in favour of conserving wild pollinators and cultivating resilience via beneficial relationships with wild plants and other endemic wildlife (White 2020). Finally, many proponents of regenerative agriculture, though not all, also argue for explicit societal goals, such as safer and more equitable labour practices, improved community health, and food sovereignty (Ikerd 2021).
Elaborating “regeneration” within the context of marine systems would sensibly begin with an identification of the appropriate aquatic analogues to soil and terrestrial nutrient cycling. Marine nutrient cycles operate at very different spatial and temporal scales than terrestrial cycles. While microbial decomposition and nutrient recycling in soils are relatively spatially discrete and happen at the scale of individual plants, in a marine setting, nutrient cycling is much more diffuse. While some amount of nutrient cycling happens in the inshore region, much decomposition and consumption of organic matter occurs in the deep ocean, meaning that these nutrients are only available for primary production in the euphotic, or surface zone, if they are returned to the surface via upwelling.
Anadromous fisheries offer another spatial complication for nutrient cycling. Fish such as salmon and alewives rely significantly on marine-derived nutrients, spending much of their life at sea feeding and growing. When they return to their natal waters to spawn and die, they transport these nutrients upriver to terrestrial ecosystems (Walters et al. 2009; Walsh et al. 2020). Anadromous fish are just one of multiple documented examples in which the fate of marine nutrients extends far from the locale of primary production (Vander Zanden et al. 2012), some of which also involve active intervention by human activities (Mathews and Turner 2017).
Thus, while a farmer can directly observe and intervene in a relatively discrete nutrient cycle that happens within the scale of their own land and practices, the marine nutrient cycle is much more diffuse, ostensibly extending well outside the possible influence of individual fishers or even entire fleets. To put this in ecological terms, when farmers enact practices to build and conserve soil, they are conserving bottom-up ecological controls at a very localized level of organization. By comparison, most decisions made in fisheries focus on specific target species and thus contribute to top-down processes that can have much more far-ranging impacts, e.g., through harvests of large predators and omnivores (Baum and Worm 2009).
Ecosystem controls: top-down, bottom-up, and wasp-waist
Absent straight-forward analogues for soil, an alternative set of analogies for thinking about regenerative seafood can be found in agroecology's emphasis on cultivating diversified agroecosystems that are intentionally integrated with wild biodiversity. Regenerative practices seek to emulate, or replace, critical controls in ecosystems, specifically those that seek to foster diversity and resilience (Pereira et al. 2018). Industrial agricultural practices simplify production ecosystems to maximize individual commodity crops, but agroecology instead seeks to maximize on-farm biodiversity, both cultivated and wild, to produce a portfolio of foods while also building resilience and creating space for regenerative processes to thrive. This is similar to the philosophy of ecosystem-based fisheries management (EBM), which seeks to scale out from a singular focus on target commodity species to conserve and rebuild assemblages of species that are connected via habitat and food webs (Pikitch et al. 2004). However, while much EBM in practice has retained a focus or bias toward specific target species, there are also examples here that take a whole systems approach and emphasize the movement of energy and nutrients through the system (Dolan et al. 2016).
Attention to the direction of ecosystem controls in fisheries, e.g., top-down and bottom-up, may also be key to thinking about regeneration in fisheries and other marine foods. While fishing itself often functions as a top-down control (Baum and Worm 2009); other patterns matter in marine food webs, including bottom-up controls, where the small drive the large via food supply, and wasp-waist controls, where the middle (e.g., small pelagic fish) controls both the large and the small (Cury et al. 2003). With respect to the latter, the contributions of forage fish to the productivity of commercialized fisheries have been shown to be noteworthy (Pikitch et al. 2014), suggesting that an emphasis on wasp-waist dynamics may be an effective policy entry point for management actions that conserve regenerative ecological relationships. Importantly, regional biogeographic variation plays an important role in determining which of the three patterns of controls—top-down, bottom-up, or wasp-waist—dominates specific community structures, e.g., the proximity to upwellings (Cury et al. 2000). There is also the potential for multiple stable states—where histories of environmental change or human exploitation have pushed specific marine communities from one control regime and food web assemblage to a second, perhaps depleted or coerced state (Cury et al. 2000).
Atlantic lobster fisheries provide a robust example of an anthropogenically coerced stable state—where the system shows signs of long-term sustainability but fails to achieve self-organization, and therefore requires constant management and subsidies to maintain the desired state (Angeler et al. 2020). As noted, these fisheries have long been hailed as sustainability success stories, but research has shown that fishers' long-established practices that favour the conservation of breeding lobsters have turned the Gulf of Maine ecosystem into a veritable lobster monoculture (Steneck et al. 2011). Top predators are generally absent, and overall diversity is low (Steneck and Wahle 2013). Moreover, the lobster populations have become so large that they rely on significant food subsidies in the form of baitfish (Grabowski et al. 2010). The predicament is remarkably similar to that seen in industrial agroecosystems, where soils and landscapes are devoid of diversity, lacking in resilience to pests and environmental change, and heavily reliant on nutrient subsidies in the form of petroleum-based fertilizers (Rist et al. 2014).
From coerced to regenerative systems
Coerced systems such as Atlantic lobster can be considered sustainable, at least for a time, but they become increasingly brittle the longer they are coerced (Angeler et al. 2020; Heilpern et al. 2022). To understand why, it is helpful to consider coerced systems within a broader organizational framework that contrasts them with other possible patterns of outcomes, including regenerative outcomes (Fig. 1). Elsewhere, I argue that the regenerative capacity of systems can be understood as an interplay among two key organizing principles—flexibility and diversity—that together establish four archetypes or categories of systems: regenerative, degenerative, coerced, and impoverished (Loring 2021). Because this framework focuses on the organization of the food system rather than the specific practices, technologies, or ecological resources being cultivated and harvested, it is as applicable to aquatic-based food systems as it is to terrestrial ones.
Fig. 1.
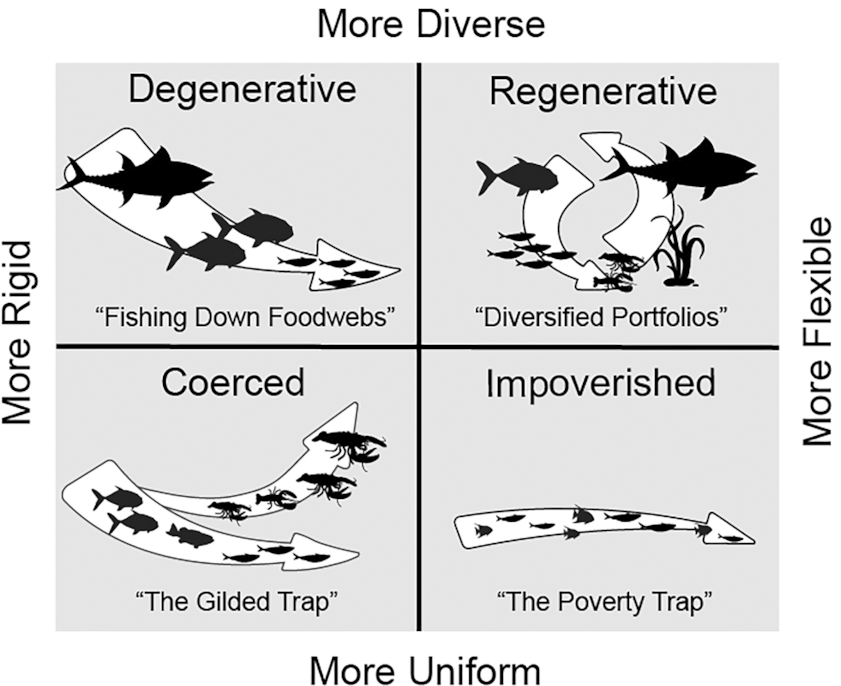
Of the four, degenerative systems—cases of fishing down or through food webs—have been extensively discussed in fisheries (Pauly et al. 1998; Essington et al. 2006). They occur where economic priorities fixate on specific, highly valued resources, and human use and management are too inflexible to switch or diversify when the targeted species are in decline. Only when a species collapses are fishers forced to switch to the next resource in line. Coerced systems, as described above, are like degenerative systems in that the economic focus is relatively inflexible and focused on single, high-value resources, but actions are taken to subsidize that resource via external inputs at the expense of ecosystem resilience. The third type, impoverished systems, often exist as the legacy of past degenerative or coerced systems. That is, these are systems where a previous regime of fishing caused significant collapse, leaving ecosystems and also human communities in a poverty trap (Cinner 2011). Newfoundland cod fisheries provide a lasting and well-known example of such a system, where neither the ecological communities nor human communities have returned to anything similar to a pre-collapse state despite many attempts to make such a recovery possible (Schlüter et al. 2019; Sguotti et al. 2019).
The fourth and final type, regenerative systems, are those where harvesters’ priorities are flexible and highly responsive to the population dynamics of multiple species within the ecological community. To my knowledge, no commercial fisheries have been examined and identified from this specific framing, though research on coral reef fisheries has shown the potential for achieving win-win outcomes for both fisheries and biodiversity (Cinner et al. 2016). Indigenous fisheries also offer a notable example, as traditional subsistence systems for many Indigenous cultures incorporate fish as just one part of a broader portfolio of aquatic and terrestrial foods and a strategy of switching among multiple options in response to changes in distribution and abundance (Berkes 2010; Loring and Gerlach 2010).
Pathways to regenerative fisheries
To move fisheries toward more regenerative configurations, it is necessary to first develop a critical understanding of their current state, specifically regarding which of the four regimes described above any specific fishery presently falls into. As noted, the state of many global fisheries is contested. Some have made broad declarations regarding the sustainability of multiple fisheries around the world (Hilborn et al. 2020), but ongoing global declines in marine biodiversity and biomass raise a question of whether the regenerative processes that support these fisheries are intact or if some fisheries only appear sustainable because, as with Atlantic lobster, they are being coerced by practices that are slowly remaking and simplifying the marine ecosystems that support them.
Different approaches will likely be necessary to steward regenerative processes in marine ecosystems depending on the nature of the fishery and the dominant ecological controls at play from one system to the next. There are clearly critical differences between the ecologies of aquaculture, sedentary fisheries, and wild-caught fin fisheries, and understanding these will be critical for designing and deploying effective and appropriately scaled institutions.
Indigenous clam gardens along the northwest Pacific Coast of North America are one example of a sedentary mariculture system that exhibits a regenerative pattern. These gardens are beach areas that people have constructed or modified with a rock wall located near the low-tide line. High tides deposit sand and other substrate on the shoreward side of the wall, increasing the amount of habitat available for clams and other species (Lepofsky et al. 2020). Historically, clam gardens are a technology used to diversify and increase food production, one that historically scaled with population growth while not causing environmental degradation (Holmes et al. 2022). While they have some features of a coerced system, for example, the effort taken to create and maintain the rock walls to favour a specific food resource, the gardens have been shown to not only increase clam productivity but also contribute positively to adjacent biodiversity and ecosystem structure (Cox et al. 2019; Cox 2021).
By comparison, strategies to promote regeneration in fin fisheries will no doubt look very different. Even among fin fisheries, there are dramatic differences among the ecologies of anadromous, small pelagic, and highly migratory species, and these differences will no doubt require different kinds of institutions operating at appropriate societal levels, from highly localized approaches to expansive international agreements. In settings where wasp-waist controls play a significant role in overall ecosystem structure and function, for example, forage fish may be a critical indicator species for understanding how to best structure a fishing portfolio that works with rather than against wasp-waist dynamics. In other settings, attention to predator species may be more appropriate if ecosystems have coevolved around top-down controls, e.g., keystone species, for rebuilding impoverished marine ecosystems. There is likewise evidence that protected areas and strategic closures enhance fisheries biomass while not necessarily disrupting fisheries, which may signal that spatial management approaches are effective at promoting ecosystem-wide regeneration (Di Lorenzo et al. 2016; Cinner et al. 2019).
Cinner and colleagues (2019) report on one such system: a customary system of rotational closures in reef fisheries in Papua New Guinea that enhances the biomass of fished species. The system is similar in function to fallow practices in agriculture, in that the closures afford the ecological community time to recover from fishing pressure. The authors note, however, that despite the periodic short-term biomass gains provided by the closures, data for a 16-year period suggests that overall fisheries biomass is declining. This could be interpreted as evidence of coercion, or that fishing pressure remains too high for the customary fallow system to mitigate. The latter has been observed in swidden agricultural systems, when pressures from markets and demographic change push farmers to shorten their fallows and flip the practice from regenerative to degenerative in nature (Kleinman et al. 1995).
Self-organization is an important difference between coerced and regenerative systems that may be a critical benchmark for disentangling the two. As noted earlier, Angeler et al. (2020) argue that coerced systems are those that fail to self-organize, and instead achieve some level of homeostasis via the application of ad hoc subsidies and other input-intensive human interventions that prop up specific, highly valued resources. A self-organized system, by comparison, is one that has developed through a mix of agency, adaptation, and selection, with sufficient social and ecological feedbacks to maintain some level of stability in the face of change (Lansing 2003). Generally, these feedbacks entail both important ecological relationships as well as flexible and highly adapted social institutions capable of learning from and responding to change. Self-organization is evident in the examples of clam gardens and reef fisheries above, in that both are deeply embedded in local social and cultural systems. However, there are limits to the resilience to external drivers of change that self-organization provides: colonialism drove the collapse and abandonment of clam gardening, and contemporary societal demands on reef fisheries appear to be pushing the customary system of closures past its regenerative potential.
While the above two examples are traditional and small-scale in nature, this final point about the role of exogenous social and economic pressures is critical to commercially developed fisheries as well. Market forces can lag behind ecological changes, and fishers do not always respond in straightforward ways to economic incentives in multispecies fisheries (Birkenbach et al. 2020). Consumer seafood purchasing behaviour has been shown to change relatively quickly as supply changes (Stoll et al. 2021), so human dimensions research could explore how best to govern and incentivize fisheries and arrange value chains such that fishers can respond quickly to ecological feedback by switching to alternative species while not sacrificing market access or livelihood stability. As with agroecology's inherent attention to justice, a regenerative food system framing for fisheries may align well with the needs and issues facing small-scale fisheries (Short et al. 2021). In all cases, it is clear that the broader socioeconomic system in which fisheries are managed ought to approach them with a level of flexibility that works with both human needs and critical ecological patterns and processes.
A word of caution is important here regarding greenwashing. Already in agribusiness we are seeing numerous questionable proclamations and commitments to regenerative practices with little transparency or clarity regarding how outcomes can be measured and monitored (Tittonell et al. 2022). Given that, as explained above, marine nutrient cycles and successional processes are far more geographically diffuse and difficult to monitor, any claims regarding the regenerative qualities of specific fisheries or other marine resource harvests should be considered with a high degree of skepticism. More science, including at the policy–practice interface, is necessary to fully explore the coherence of regenerative system design in a marine context.
All of this is perhaps yet another rationale for reorienting how we approach fisheries, from treating them as resources to acknowledging their critical and complex role as food (McClanahan et al. 2013; Levkoe et al. 2017). A next step would be to reconsider fisheries case studies in a way that links explicit modeling of the dominant ecological controls in marine food webs with the social and market factors driving harvest and enabling or limiting harvest diversification and flexibility. Just as there may be cases of sustainable fisheries that are in fact coerced and in need of re-evaluation, there may also be fisheries regimes (e.g., small-scale fisheries and alternative seafood networks) that are successfully making space for ecological regeneration while also providing food and economic services to society. At a minimum, it is clear that we need to address the market fetishization of high-value commodity species, as this culture of fishing creates strong economic incentives against moving toward regeneration.
Acknowledgments
This work was supported in part by the Arrell Food Institute at the University of Guelph.
References
Acheson J.M. 1990. The lobster fiefs revisited. In Question of the commons. Edited by B.J. McCay, J.L. Anderson. The University of Arizona Press, Tuscon, AZ.
Angeler D., Chaffin B.C., Sundstrom S.M., Garmestani A., Pope K.L., Uden D.R., et al. 2020. Coerced regimes: management challenges in the Anthropocene. 25. Available from https://pub.epsilon.slu.se/16709/ [accessed 28 October 2020].
Barrett L.T., Swearer S.E., Dempster T. 2019. Impacts of marine and freshwater aquaculture on wildlife: a global meta-analysis. Reviews in Aquaculture, 11(4): 1022–1044.
Baum J.K., Worm B. 2009. Cascading top-down effects of changing oceanic predator abundances. Journal of Animal Ecology, 78(4): 699–714.
Berkes F. 2010. Shifting perspectives on resource management: resilience and the reconceptualization of “natural resources” and “management”. Mast, 9(1): 13–40.
Birkenbach A.M., Cojocaru A.L., Asche F., Guttormsen A.G., Smith M.D. 2020. Seasonal harvest patterns in multispecies fisheries. Environmental and Resource Economics, 75(3): 631–655.
Cheney J. 2021. Trying to find answers for the mysterious Bering Sea crab collapse. Sustainable Fisheries UW, 16 November. Available from https://sustainablefisheries-uw.org/mysterious-bering-sea-crab-collapse/ [accessed 12 October 2022].
Cinner J.E. 2011. Social-ecological traps in reef fisheries. Global Environmental Change.
Cinner J.E., Huchery C., Macneil M.A., Graham N.A.J., Mcclanahan T.R., Maina J., et al. 2016. Bright spots among the world's coral reefs. Nature, 535(7612): 416–419.
Cinner J.E., Lau J.D., Bauman A.G., Feary D.A., Januchowski-Hartley F.A., Rojas C.A., et al. 2019. Sixteen years of social and ecological dynamics reveal challenges and opportunities for adaptive management in sustaining the commons. Proceedings of the National Academy of Sciences, 116(52): 26474–26483.
Costello C., Ovando D., Clavelle T., Strauss C.K., Hilborn R., Melnychuk M.C., et al. 2016. Global fishery prospects under contrasting management regimes. Proceedings of the National Academy of Sciences, 201520420.
Cox K.D., Gerwing T.G., Macdonald T., Hessing-Lewis M., Millard-Martin B., Command R.J., et al. 2019. Infaunal community responses to ancient clam gardens. ICES Journal of Marine Science, 76(7): 2362–2373.
Cox K.D. 2021. Intertidal resource cultivation over millennia structures coastal biodiversity. Thesis. Available from https://dspace.library.uvic.ca/handle/1828/13638 [accessed 6 April 2023].
Cury P. 2000. Small pelagics in upwelling systems: patterns of interaction and structural changes in “wasp-waist” ecosystems. ICES Journal of Marine Science, 57(3): 603–618.
Cury P., Shannon L., Shin Y.-.J. 2003. The functioning of marine ecosystems: a fisheries perspective. In Responsible fisheries in the marine ecosystem. pp. 103–123.
Di Lorenzo M., Claudet J., Guidetti P. 2016. Spillover from marine protected areas to adjacent fisheries has an ecological and a fishery component. Journal for Nature Conservation, 32: 62–66.
Dolan T.E., Patrick W.S., Link J.S. 2016. Delineating the continuum of marine ecosystem-based management: a US fisheries reference point perspective. ICES Journal of Marine Science, 73(4): 1042–1050.
Essington T.E., Beaudreau A.H., Wiedenmann J. 2006. Fishing through marine food webs. Proceedings of the National Academy of Sciences of the United States of America, 103(9): 3171–3175.
FAO. 2022. The state of world fisheries and aquaculture 2022. Towards blue transformation. FAO, Rome, Italy.
Farmery A.K., Allison E.H., Andrew N.L., Troell M., Voyer M., Campbell B., et al. 2021. Blind spots in visions of a “blue economy” could undermine the ocean's contribution to eliminating hunger and malnutrition. One Earth, 4(1): 28–38.
Gaupp F., Ruggeri Laderchi C., Lotze-Campen H., Declerck F., Bodirsky B.L., Lowder S., et al. 2021. Food system development pathways for healthy, nature-positive and inclusive food systems. Nature Food, 2(12): 928–934.
Gentry R.R., Alleway H.K., Bishop M.J., Gillies C.L., Waters T., Jones R. 2020. Exploring the potential for marine aquaculture to contribute to ecosystem services. Reviews in Aquaculture, 12(2): 499–512.
Grabowski J.H., Clesceri E.J., Baukus A.J., Gaudette J., Weber M., Yund P.O. 2010. Use of herring bait to farm lobsters in the Gulf of Maine. PLoS One, 5(4): e10188.
Heilpern S.A., Sethi S.A., Barthem R.B., Batista V.Da.S., Doria C.R.C., Duponchelle F., et al. 2022. Biodiversity underpins fisheries resilience to exploitation in the Amazon river basin. Proceedings of the Royal Society B: Biological Sciences, 289(1976): 20220726.
Hilborn R., Amoroso R.O., Anderson C.M., Baum J.K., Branch T.A., Costello C., et al. 2020. Effective fisheries management instrumental in improving fish stock status. Proceedings of the National Academy of Sciences, 117(4): 2218–2224.
Holmes K., Lepofsky D., Smith N.F., Crowell T.D., Salomon A.K. 2022. Ancestral sea gardens supported human settlements for at least 3,800 years on the Northwest Coast of North America. Frontiers in Earth Science, 10. Available from https://www.frontiersin.org/articles/ [accessed 6 April 2023].
Ikerd J. 2021. The economic pamphleteer: realities of regenerative agriculture. Journal of Agriculture, Food Systems, and Community Development, 10(2): 1–4.
Jacquet J., Pauly D. 2022. Reimagining sustainable fisheries. PLoS Biology, 20(10): e3001829.
Kleinman P.J.A., Pimentel D., Bryant R.B. 1995. The ecological sustainability of slash-and-burn agriculture. Agriculture, Ecosystems & Environment, 52(2–3): 235–249.
Koehn J.Z., Allison E.H., Golden C.D., Hilborn R. 2022. The role of seafood in sustainable diets. Environmental Research Letters, 17(3): 035003.
Lansing J.S. 2003. Complex adaptive systems. Annual Review of Anthropology, 32(1): 183–204.
Lepofsky D., Toniello G., Earnshaw J., Roberts C., Wilson L., Rowell K., Holmes K. 2020. Ancient anthropogenic clam gardens of the Northwest Coast Expand Clam Habitat. Ecosystems.
Levkoe C.Z., Lowitt K., Nelson C. 2017. “Fish as food”: exploring a food sovereignty approach to small-scale fisheries. Marine Policy, 85: 65–70.
Loring P.A., Gerlach S.C. 2010. Outpost gardening in interior Alaska: food system innovation and the Alaska native gardens of the 1930s through the 1970s. Ethnohistory, 57(2): 183–199.
Loring P.A. 2021. Regenerative food systems and the conservation of change. Agriculture and Human Values, 1–13.
Lovelock C.E., Duarte C.M. 2019. Dimensions of blue carbon and emerging perspectives. Biology Letters, 15(3): 20180781.
Mathews D.L., Turner N.J. 2017. Chapter 9 – ocean cultures: northwest coast ecosystems and indigenous management systems. In Conservation for the anthropocene ocean. Edited by P.S. Levin, M.R. Poe. Academic Press. pp. 169–206.
Mcclanahan T., Allison E.H., Cinner J.E. 2013. Managing fisheries for human and food security. Fish and Fisheries, 16(1): 78–103.
Newton P., Civita N., Frankel-Goldwater L., Bartel K., Johns C. 2020. What is regenerative agriculture? A review of scholar and practitioner definitions based on processes and outcomes. Frontiers in Sustainable Food Systems, 4.
Palomares M.L.D., Froese R., Derrick B., Meeuwig J.J., Nöel S.-.L., Tsui G., et al. 2020. Fishery biomass trends of exploited fish populations in marine ecoregions, climatic zones and ocean basins. Estuarine, Coastal and Shelf Science, 243: 106896.
Pauly D., Christensen V., Dalsgaard J., Froese R., Torres F. 1998. Fishing down marine food webs. Science, 279(5352): 860–863.
Pearson C.J. 2007. Regenerative, semiclosed systems: a priority for twenty-first-century agriculture. BioScience, 57(5): 409–418.
Pereira L., Wynberg R., Reis Y. 2018. Agroecology: the future of sustainable farming? Environment: Science and Policy for Sustainable Development, 60(4): 4–17.
Pikitch E.K., Santora C., Babcock E.A., Bakun A., Bonfil R., Conover D.O., et al. 2004. Ecosystem-based fishery management. Science, 305(5682): 346–347.
Pikitch E.K., Rountos K.J., Essington T.E., Santora C., Pauly D., Watson R., et al. 2014. The global contribution of forage fish to marine fisheries and ecosystems. Fish and Fisheries, 15(1): 43–64.
Rhodes C.J. 2017. The imperative for regenerative agriculture. Science progress, 100(1): 80–129.
Rist L., Felton A., Nyström M., Troell M., Sponseller R.A., Bengtsson J., et al. 2014. Applying resilience thinking to production ecosystems. Ecosphere, 5(6): 1–11.
Schlüter M., Haider L.J., Lade S.J., Lindkvist E., Martin R., Orach K., et al. 2019. Capturing emergent phenomena in social-ecological systems. Ecology and Society, 24(3).
Sguotti C., Otto S.A., Frelat R., Langbehn T.J., Ryberg M.P., Lindegren M., et al. 2019. Catastrophic dynamics limit Atlantic cod recovery. Proceedings of the Royal Society B: Biological Sciences, 286(1898): 20182877.
Short R.E., Gelcich S., Little D.C., Micheli F., Allison E.H., Basurto X., et al. 2021. Harnessing the diversity of small-scale actors is key to the future of aquatic food systems. Nature Food, 2(9): 733–741.
Steneck R.S., Hughes T.P., Cinner J.E., Adger W.N., Arnold S.N., Berkes F., et al. 2011. Creation of a gilded trap by the high economic value of the maine lobster fishery. Conservation Biology, 25(5): 904–912.
Steneck R.S., Wahle R.A. 2013. American lobster dynamics in a brave new ocean. Canadian Journal of Fisheries and Aquatic Sciences, 70(11): 1612–1624.
Stoll J.S., Harrison H.L., De Sousa E., Callaway D., Collier M., Harrell K., et al. 2021. Alternative seafood networks during COVID-19: implications for resilience and sustainability. Frontiers in Sustainable Food Systems, 5: 614368.
Tittonell P., El Mujtar V., Felix G., Kebede Y., Laborda L., Luján Soto R., De Vente J. 2022. Regenerative agriculture—agroecology without politics? Frontiers in Sustainable Food Systems, 6. Available from https://www.frontiersin.org/articles/ [accessed 23 November 2022].
Vander Zanden H.B., Bjorndal K.A., Inglett P.W., Bolten A.B. 2012. Marine-derived nutrients from green turtle nests subsidize terrestrial beach ecosystems. Biotropica, 44(3): 294–301.
Walsh J.C., Pendray J.E., Godwin S.C., Artelle K.A., Kindsvater H.K., Field R.D., et al. 2020. Relationships between Pacific salmon and aquatic and terrestrial ecosystems: implications for ecosystem-based management. Ecology, 101(9): e03060.
Walters A.W., Barnes R.T., Post D.M. 2009. Anadromous alewives (Alosa pseudoharengus) contribute marine-derived nutrients to coastal stream food webs. Canadian Journal of Fisheries and Aquatic Sciences, 66(3): 439–448.
White C. 2020. Why regenerative agriculture? The American Journal of Economics and Sociology, 79(3): 799–812.
Information & Authors
Information
Published In

FACETS
Volume 8 • January 2023
Pages: 1 - 6
Editor: S.J. Cooke
History
Received: 20 January 2023
Accepted: 25 April 2023
Version of record online: 24 July 2023
Copyright
© 2023 The Author. This work is licensed under a Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Data Availability Statement
Not applicable.
Key Words
Sections
Subjects
Plain Language Summary
Can fisheries and other aquatic foods be “regenerative”?
Authors
Author Contributions
Conceptualization: PAL
Formal analysis: PAL
Visualization: PAL
Writing – original draft: PAL
Competing Interests
The authors declare there are no competing interests.
Metrics & Citations
Metrics
Other Metrics
Citations
Cite As
Philip A. Loring. 2023. Can fisheries be “regenerative”? Adapting agroecological concepts for fisheries and the blue economy. FACETS.
8(): 1-6. https://doi.org/10.1139/facets-2023-0011
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
There are no citations for this item